[回上頁 | 顯示此文件為可列印格式]
此文件提供者: shuYing - [評分 : 0.00 (0 票選) | 評分!]
OVERALL REVIEW OF GENETICAL
RESEARCH ON CHINESE PTGS IN FRANCE AND PROSPECTS FOR EXPLOITING CHINESE
CROSSBRED ANIMALS
C. LRGAULT AND J.P.BIDANEL
Institut National de la Recherche
Agronomique
Station de Genetique quantitative et
appliquee
78352 Jouy en Josas Cedex – FRANCE
SUMMITRY
The present review summarizes the
main results obtained in France over 12 years of research on three native breeds
of pigs from the People’s Republic of China: Meishan (MS), Jiax-ing (JX) and
Jinhua (JH). The first part is devoted to breed evaluation regarding
reproduction, growth, feed efficiency, body composition, meat quality, behaviour
and resistance lo diseases. On the basis of an estimation of Dickerson’s
crossbreeding parameters between Meishan and Large White breeds, the second part
of the review presents and discusses several strategies for taking advantage of
prolific Chinese breeds as maternal components in crossbreeding schemes under
different production systems.
I -INTRODUCTION
A limited number of native breeds
of the People's Republic of China exhibits exceptional reproductive ability.
They could then be incorporated as a component of maternal lines in European
breeding programs in order to improve sow numerical productivity. That is why
breeding animals from three native Chinese breeds of pigs: Meishan (MS), Jiaxing
(JX) and Jinhua (JH) were introduced into France in 1979. Later, Chinese pigs
have also been imported by several other countries: Hungary, Japan, the
Netherlands, Great-Britain, the USA,….
Chinese pigs represent a valuable
resource for basic research in many fields of biology and pig industry including
reproduction, growth, body composition, meat quality, nutrition, resistance to
deseases and behaviour. Several research programmes have been set up in France
in order to evaluate these three lines and to assess their economic value as a
component of the maternal genetic type in various crossbreeding systems. Due to
the extremely narrow genetic base of these lines, any extrapolation to the whole
breeds is of course not advisable. Nevertheless, these programmes have provided
useful results concerning their performance under various production systems and
their physiological characteristics.
The first purpose of the present
review is to evaluate these three local breeds in comparison with usual European
breeds such as Large White (LW) and French Landrace (LF) on the basis of the
main results obtained in France over the last 12 years. The second purpose is to
define and discuss the different strategies of utilisation of these breeds under
various production and crossbreeding systems. Most of these results have already
been widely discussed during the First Symposium on Chinese pigs held Toulouse
in July 1990.
Source: Proceedings of the
International Symposium on Chinese Pig Breeds in Harbin, China. 1992. pp. 10-24.
II. BREED EVALUATION
A first review of experimental
results obtained on Chinese Pigs in France has already been published (Bidanel
et al., 1990a; 1990b; 199]a). In the present review, the emphasis will be placed
un recent development of these investigations.
A-Reproduction
The
three Chinese lines reach puberty very early. Mean age at first oestrus was 81,
9l and 109 days in MS, JX and JH, respectively in the first study of Legault and
Caritez (1983). These observations were recently confirmed on a larger scale
study involving MS and LW breeds (Despres et al. 1992; figure I).
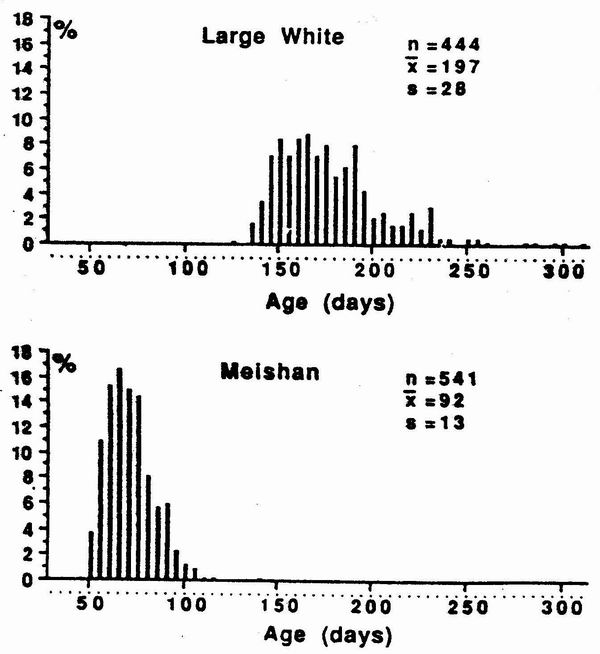
Figure 1. Distribution of age at puberty (days ) in Large While and
Meishan gilts
(Dcspres et al 1992). |
Compared to LW animals, MS
pigs are characterized by an earlier growth pattern of1he reproductive tract in
females, accessory glands in males and gonads in both sexes (Prunier et al.
1987). This low age at puberty could result from an earlier start in sexual
development due to an increase in hypothalamic and pituitary secretions and also
from the lack of a waiting phase in the ovary development generally observed in
European gilts in the weeks preceeding puberty (Prunier et al., 1987).
Fl Chinese x European gills
also reach puberty very early, between 85 and 110 days of age under normal
temperate climate (Legault and Caritez, 1983) and between 136 and 150 days of
age under tropical climate (Canope et al. 1990).
An experiment was designed to
estimate the effects of an early age at first farrowing on body development,
longevity and productivity of sows (Gauthier et al. 1992). Sixty-one Fl French
Landrace x Meishan crossbred gills (LF ´ MS) were mated either at 5 (MS) or at 7
months of age (M7) and compared over five successive parities to 29 control F1
Finch Landrace x Large White crossbred gilts (LF ´ LW) mated at 7 months of age.
The highest longevity as well as the best reproductive performance were observed
in LF ´ MS sows mated at 5 months. The delay in body development observed over
the first reproductive cycle became non significant in the second and later
parities. The numerical productivity estimated over the periapt from 150 days of
age to the weaning of the fifth litter was 29.6, 26.7 and 23.4 pigs weaned / sow
/year for M5, M7 and control sow', respectively. The excellent longevity of Fl
sows as compared to herd-contemporaries or the French national reference is
illustrated by figure 2a under. temperate climate (Gauthier et al., 1992) and by
figure 2b under tropical climate (Delate et al 1991).
|
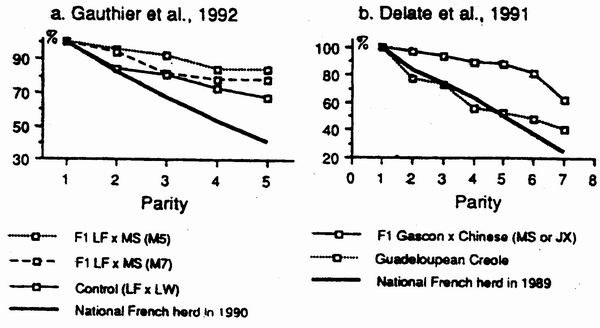
Figure 2. Comparison of longevity of Chinese
crossbred sows to contemporaries
under temperate and tropical climate. |
The high litter size of MS gilts
is mainly due to a low embryonic mortality, their ovulation rate being
comparable at the same age and inferior at the same oestrus number with those of
LW gilts (Bolet et al., 1986; Bazer et al. 1988a, Bidanel et al. 1990c). The
physiological mechanisms underlying the high conceptus survival of MS gilts are
discussed by Terqui et al. (1992).
In the French experiment, MS
females reached the same high levels of prolificacy as those obtained in China,
with a superiority over LW or LF of about three piglets born per litter (Legault
and Caritez, 1983; Bidanel et al., 1989). On the other hand, JX and JH French
lines do not appear superior to European contemporaries, at least in the first
three parities (figure 3). After the third parity, JX saws farrow larger litters
than LW or JH, but remain at a lower level than MS sows.
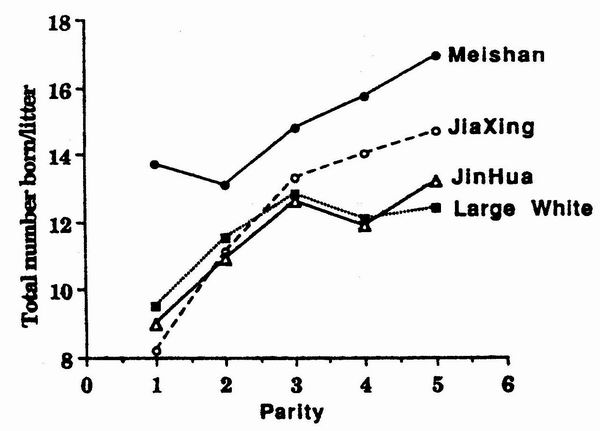
Figure 3. Comparative variation of litter size at birth (total
number born) according to
parity in Lange White, Jinhua, Jiaxing and Mcishan sows. |
The aim of a recent study (Mandonnet et al.,
1992) was to test the hypothesis of a major gene contributing to litter size in
pigs, using all available data on pure MS and crossbred MS
´ LW sows. The total number. of piglets born and the number of
piglets born alive per litter ware recorded in 1903 and 1660 litters,
respectively. Prolificacy was considered as a trait of the dam. Under the
hypothesis of a major locus with fixed A and B alleles in MS and LW breeds, we
were expecting an AB homogeneous Fl population and backcross and F2 populations
in which the 2 alleles were segregating: 1/2 AB and 1/2 AA (or BB) for the
backcrosses, 1/4AA, 1/2AB and 1/4BB for the F2. This hypothesis was tested
versus a sporadic inheritance by use of a segregation analysis method. Non
significant likelihood ratios led us to conclude that prolificacy differences
between MS and LW breeds could not be explained by one major gene and that the
high prolificacy in MS was probably due to a very long term selection in favour
of litter size.
MS and to a lesser extent JX sows also have
excellent maternal abilities, as shown by the large litters they are able to
foster up to weaning (Depres et al., 1991). After adjusting for number born
alive, MS and JX exhibit an advantage in survival rate over LW of 5 and 3%,
respectively (Bidanel et al.,1990a). These good mothering abilities are partly
due to behavioural characteristics (Meunier–Salaun et al., 199l), but also to
the high number of teals in both breeds: 16.1 in MS, 18.8 in JX (Bidanel and
Legault, 1986). Nevertheless, MS have a lower milk production than LW sows (Legault
and Caritez, 1983; Bidanel et al., 1989). Weaning–oestrus interval (5.2+ 0.8 vs
7.5+ 0.9 days) as well as weaning fertilization interval(12.4+ 1.8 vs 12.4+ 1.5
days) were reduced in MS as compared to LW saws (Despres et al.,1992).
B-Growth and feed efficiency
MS, JX and above all JH are much lighter at
birth than LW (between 900g and 1000g for MS, around S50g for JX and 650 g for
JH vs. 1100-1300g for LW-Legault et al., 1982; 3idanel et al., 1990d; Le
Dividich tt al., 1991). MS piglets have a greater percentage of body dry matter
and protein, are more mature and have higher (+75%) plasma concentrations of
free fatty acids than LW. This would explain their better resistance to cold (Le
Dividich et al., 1991). The size disadvantage of MS, JX and JH with regard to LW
keeps on increasing all through growth (figure 4). It reaches 26, 32 and 41 kg
respectively at 154 days, 58,100 and 120 kg at first farrowing and 80, 130 and
150 kg at adult weight.
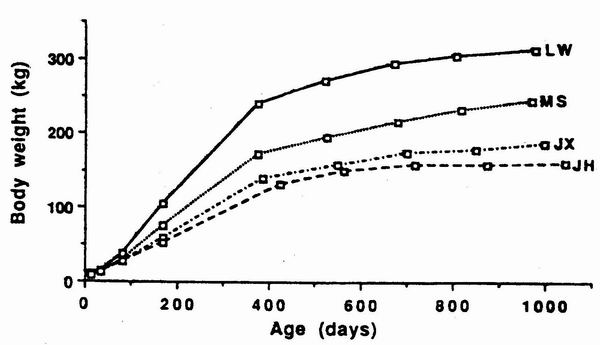
Figure 4. Growth pattcrn of Large White I.W), Mcishan (MS), Jiaxing
(JX), and
Jinhua (JH) fcmalcs (Bidancl et al., 1990a). |
Feed efficiency of purebred Chinese animals
has so far not been measured over the whole post – weaning period.
Extrapolations from the performance of crossbred pigs lead to a respective
disadvantage of 0.9, 1.4 and 1.6 points of food conversion ratio for MS, JX and
JH, respectively, compared to European controls (Legault et at., 1985). However,
he real difference is certainly larger, as iced efficiency presents
non-negligible heterosis effects. The ability of MS pigs to digest a fibrous
diet was studied by Fevrier et al. (1988). It was found that MS digest a fibrous
diet more efficiently than LW, but retain less nitrogen.
Feed intake is similar in LW and MS in young
animals (up to 30 kg live weight). After-wards, the increase with weight in feed
intake is lower in MS animals (Bonneau et al., 1990). The difference in growth
rate and body composition between the two breeds is mainly associated with a
much lower daily protein and muscle deposition in MS animals.
C-Body composition and meat quality
Direct estimations of body composition of MS
as compared with European breeds were made on relatively small samples by
Poilvet et al. (1990) and by Bidanel et el. (1991b). However, most of the
available results on carcass traits come from extrapolations based on
performance of Chinese crossbreds. These extrapolations are much less subject to
be criticized than those for feed efficiency, as carcass traits have an
essentially additive genetic variability. Indirect estimations of killing out
percentage give a disadvantage of 2-3 percentage points for MS, no disadvantage
for JX and an advantage of 3-4 percentage points for JH over LW (Legault et al.,
1985; Bidanel et al., 1992). Direct measurements confirm the low killing out
percentage of MS 5rrnales:-3.4 points compared with LW according to Poilvet et
al. (1990); -4.5 points compared with Pietrain according to Bidanel et al.
(1991b). The three Chinese breeds have a much worse body composition than usual
European breeds. The estimated carcass lean content reaches 29-30% for JH and
33-35% for MS and JX, against about 53-54% for LW or LF and 62% in Pi4train (Legault
et el., 1985; Bidanel et al., 199lb; 1992). It is associated with a much thicker
4<in and a higher bone weight (Poilvet et el., 1990; Bidanel et al. 199lb). The
important bone content of MS is mainly due to their high head weight (+0.72 kg
over a European crossbred control, Legault et el., l985; +1.55 kg over Pietrain,
Bidanel et al., 199lb). A similar phenomenon exists for JX, but not for JH (Legault
et el., l985).
According to Bonneau et al. (1990), the lower
potential for muscle growth of MS pig is already present at birth via a reduced
number of muscle fibers. The smeller postnatal increase in arose-sectional area
of existing type II fibers in MS pigs further enlarges the difference between
the two breeds. The more oxydative metabolism of MS muscle is observed from
birth onwards while the lower glycolytic activity is acquired during postnatal
growth. Another approach for determining the distribution of fibres consists in
measuring the distribution of lactate dehydrogenase (LDH) isozymes because LDH
is a major glycolytic enzyme and the proportion of isozyme 5 is high in
glycolytic fibres (white fibres) and low in oxydative fibres (red fibres). Such
a study conducted by Bousset et al. (1990) over six different muscles from three
genotypes (MS, LW and F1) led to the conclusion that significant but not
fundamental differences exist between genotypes, in good agreement with the
results from Bonneau et at. (1990).
Several studies have shown that MS pigs have
more intra muscular fat than LW (3.27% vs 1.91%; Poilvet et el., 1990) or than
Pietrain (3.07% vs 2.28% according to Bidanel et al., 1991b). These observations
were confirmed in various crossbred animals including MS or JX (Touraille et
al., 1989; Gandemer et al., 1992). The observed divergences in muscle fat
content were essentially due to breed variation in neutral lipids content.
Differences in polar lipids content and in fatty acid composition were much less
marked, even if signifcant deviations in favour of MS sometimes appeared.
The
technological quality of the meat tends to be enhanced with increasing proportions of MS genes in crossbred products. The strongest influence is on
meat colour. Chinese crossbreds have a significantly lower reflectance (i. e. a
darker coloured meat) that European centrols (Legault et a1., 1985; Bidanel et
al., 1992). Higher ultimate pH and water holding capacity of crossbred Chinese
were also obtained, but differences wert not significant For all muscles.
A
summary of results obtained in crossbreeding with MS is shown in figure 5 (for
average daily gain and feed conversion ratio) and figure 6 (for estimated lean
content of the carcass and meat quality index).
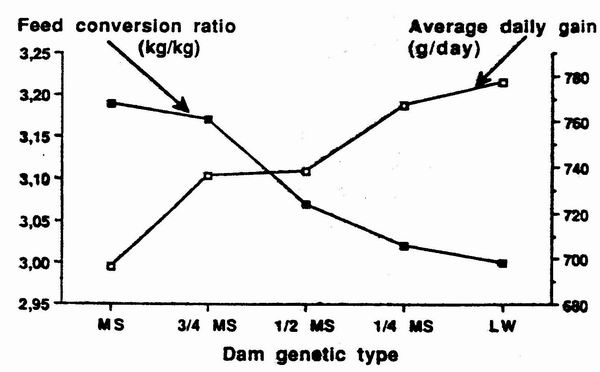
Figure 5. Variations of average daily gain and feed conversion ratio
according to the
percentage of Mcishan gencs in the darm, the sire being Pietrain
(Bidancl et al., 1992) |
Several studies clearly established the
superiority of crossbred Chinese pigs for organoleptic quality of the meat. Meat
from various 1/2 and 1 /4 Chinese crossbred pigs was judged more tender, more
juicy and more tasty than meat from purebred European pigs (LW x LF) in several
taste panel and consumer tests (Touraille et al., l989; Gandemer et al., 1992
and figure 7). This superiority seems to be highly connected with their higher
intramuscular fat content (Gandemer et al., 1992). However, the overall
acceptabili1y of the meat from crossbred Chinese pigs was not better than that
of European controls, probably due to the excessive amount of visible fat. A
superiority of Chinese crossbred over European controls animals was also
observed for dry ham sensory properties (Rousset el al., 1990).
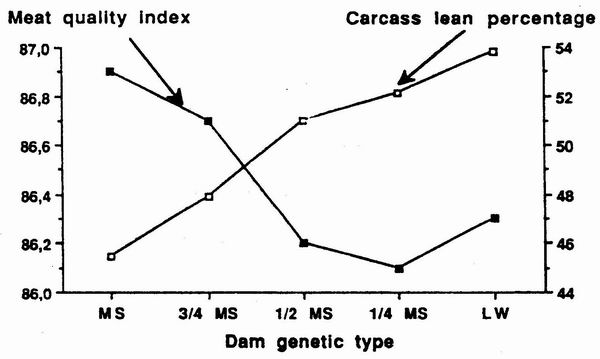
Figure 6. Variations of
meat quality index and estimated carcass lcan percentage according
to
the percentage of Mcishan genes la the dam, the sire being Pietrain
(Bidanel et al., 1992). |
D-Other aspects
A certain type of neonatal
colibaci11osis is due to the adhesion of Escherichia coli K88 on receptor sites
of the intestinal epithelium. The lack of receptor, which confers resistance to
K88 adhesion is due to a single recessive gee. A limited trial involving MS, JX
and LW breeds tend- ed to indicate that, contrary to LW, Chinese piglets lack
this receptor, thus suggesting the existence of breed variation in resistance to
this type of colibacillosis (Chappuis et al., 1984). The use of genetic
resistance to K88 colibacillosis in pig breeding schemes was discussed by
Ollivier and Renjifo (1991). In particular, the potential interest of the
linkage between K88 and transferrin loci (Bonneau et el., 1990) was highlighted.
These loci have recently been located on chromosome number 13 (Chowdhary et al.,
1991).
The general behaviour of
8-week-old piglets from the three Chinese lines was investigated through
standard tests (Mormede et al., 1984). When exposed to a new environment, MS
piglets had a lower locomotor activity and vocalized less than JX, JH and
control piglets. They also had a lower food intake latency abler 24 hours of
food deprivation (neophobia test). This behavioural reactivity had complex
relationships with the corticotropin, corticoid and epinephrine levels of the
animals. The superiority of MS sows regarding maternal behaviour as compared to
LW sows would be due to a better adaptative ability of both sows and piglets.
A comparative study of chromosomes
of Chinese (MS and JH ) and European breeds (LW and LF ) showed that the
nucleolar organising regions (NOR) were localised on both autosomal pairs no8
and 10 in Chinese pigs while the NOR were only expressed on one chromosome of
the 8th pair in most European animals (M uramatsu et al., 1990).
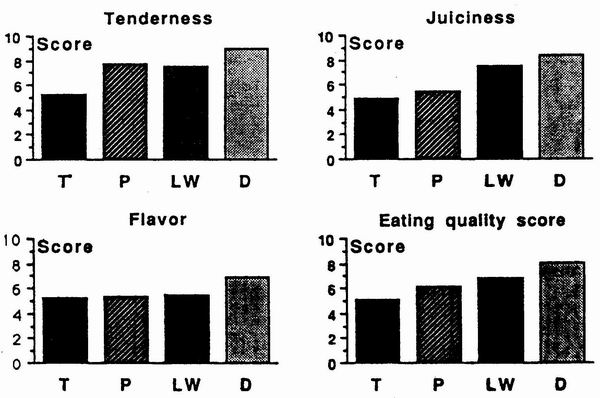
Figure 7. Effect of genetic type on eating quality of meat (Gandemer
et al., l990). |
finally, let us mention the
existence olla European cooperative project on pig gene mapping (Pig Map). The
purpose of this project which involves 16 different laboratories is to find
genetic markers which art evenly spaced and cover the whole of the pig’s genome.
Reference populations for this project are based on crosses between MS and LW
breeds in three different countries (Ollivier et a1.,1992).
Ⅲ. PROSPECTS AND DIFFICULTIES FOR EXPLOITING CHINESE BREEDS
The use of specialized sire
and dam lines has become a common practice in modern pig industry. Excellent
reproductive performance being higly appreciated in dam lines, prolific Chinese
breeds seem to be particularly recommended as maternal or grand- maternal
genotypes in breeding schemes. Unfortunately, the utilisation of this extreme
genotypes is highly compromised by their poor growth and carcass performance. A
precise knowledge of breed differences and crossbreeding parameters allows
prospective studies on the different ways of utilizing Chinese breeds in various
production systems.
In France, the performance
of MS and JX was extensively studied in both pure and crossbreeding. Dickerson's
genetic parameters (Dickerson, 1969) of the cross between MS and LW breeds were
estimated for different traits of economic interest. The main results of this
evaluation work are summarized in Table l.
Table 1 Breed differences between Mcishan (MS), Jiaxing (JX)
and Large White
(LW) breeds and beterosis effects of MS LW and JX LW crosscs (synthcsis of
French results; Bidanel, 1990).
|
|
Breed difference |
Heterosis effects |
|
|
|
direct |
maternal |
|
Item |
MS-LW |
JX-LW |
MS
´
LW |
JX
´
LW |
MS
´
LW |
JX
´
LW |
|
Age
at puberty (d) |
-101 |
(-91)b |
(-50) |
(-49) |
-c |
- |
|
Number of teats |
+3.4 |
5.9 |
(0) |
(0) |
0 |
- |
|
Piglets born alive |
+3.1 |
0.6 |
+0.9 |
- |
+2.3 |
+3.8 |
|
Piglets weaned |
+2.6 |
0.8 |
+1.2 |
- |
+2.3 |
+3.5 |
|
Adult weight (kg) |
-98 |
127 |
+27 |
- |
0 |
- |
|
Feed consumption
during lactation (kg) |
-21 |
51 |
+16 |
- |
0 |
- |
|
Average daily gain (g/d) |
-230 |
280 |
+187 |
- |
+29 |
- |
|
Food conversion ratioa |
+0.9 |
1.4 |
(0) |
- |
0 |
- |
|
Killing out %a |
-3.8 |
0.4 |
(0) |
- |
0 |
- |
|
Backfat thickness (mm)a |
+11.8 |
6.7 |
(0) |
- |
0 |
- |
|
Lean content(%)a |
-16.0 |
- |
(0) |
- |
0 |
- |
|
Ultimate pH a |
+0.12 |
- |
(0) |
- |
0 |
- |
|
Reflectance(0-l000)a |
-36 |
- |
(0) |
- |
0 |
- |
|
Imbibition time (sec.) a |
+10 |
- |
(0) |
- |
0 |
- |
a extrapolated from the performance of crossbred
pigs; b approximate value; c not estimated.
The main conclusions
of this evaluation work are as follows:
.breed differences are, at
least in MS, mainly of maternal origin for prolificacy and of direct origin for
growth traits. Grand-maternal effects are of little importance.
.both MS and JX lines
exhibit extremely large heterosis effects for reproductive and growth trails in
crosses with European breeds. As a consequence, growth rate of F1MS
´LW females is similar to that
of pure LW, their prolificacy being equivalent to that of pure MS.
.direct and maternal
epistatic recombination losses seem to be negligible. Another central step labor
an accurate comparison of various strategies involving Chinese breeds depends on
a correct prediction of their relative economic merit. Economic models must hike
into account the particularities of Chinese breeds, such as low age at puberty
and longevity, as well as their production and maintenance costs, i. e. feed
consumption, culling price and costs of selection and multiplication herds
(number of animals required, costs of nonbreeding pigs). Another important
feature concerns the genetic consequences of the high prolificacy and the early
puberty of Chinese breeds. An important increase (up to 50%-3idanel, 1988) in
the rate of response to selection can be expected by shortening the generation
interval and / or increasing selection intensities. On the other hand, the high
sexual activity of Chinese breeds strongly disturbs their growth and might lower
the efficiency of standard performance tests of young boars.
Due to the large heterosis effects
on prolificacy, the use of Chinese breeds as grand-matermal breeds (i. e. as one
of the components of the matermal genotype) is always superior to its use as a
single maternal genotype. The short-term interest of Chinese breeds greatly
depends on the relative economic impacts of production and reproduction trails,
and particularly on the part played by the pork payment system in the global
profit. For instance, the EEC market, which puts a strong emphasis on lean
weight, is much less favourable to Chinese breeds than the U. S. market, which
essentially prices pork. according to live weight. It has been clearly
demonstrated that, in France and probably in many EEC countries, there is no
conclusive short-term strategy for using MS or JX breeds, al least under the
predominant intensive production system (Legault et al., 1985; Gueblez et
a1.,1987; Bidanel, 1989).
In 1he furture, prospects for using
Chinese breeds will greatly depend on the possibilities of improving their
growth and carcass performance without impairing their reproductive merit.
Several different strategies can be proposed to reach this goal. Selecting pure
Chinese breeds for production traits could be a first solution. A 50% higher
genetic progress could be expected in pure Chinese breeds compared to
conventional breeds provided that the genetic parameters are similar and that
the sexual activity does not cause any loss in the efficiency of performance
test (Bidanel, 1989). However, the coat of this selection would be very high and
probably beyond the financial possibilities of most breeding organisations.
Creating a composite line and
selecting it for production traits appears to be a more promising solution. In
fact, the creation of a 50% Chinese
´
50% European composite line reduces by half the initial gap for production
trails. Further reductions can be expected through selection, which could also
be more efficient than in e European-type line because of an earlier puberty, a
higher prolificacy and e possible increase in the additive genetic variance (Bidanel,
1988; 19S9). On the other hand, 50% of the favourable heterosis effects on
reproduction and growth are lost (in the absence at' recombination loss). From
simulation studies, the time necessary to overtake conventional grand-maternal
lines should not exceed 3 or 4 years of intense selection for production traits.
One should add, however, the time necessary to break down linkage disequilibria
involving production and reproduction genes, so that 6 or 7 years are necessary
to reach the economic equilibrium. These theoretical predictions seem to be
confirmed by results obtained in two French companies which have undertaken the
constitution of such lines (Naveau, 1990 Looft et al. 1990). A very high
improvement of production traits was achieved without any perceptible reduction
of litter size.
Discontinuous crossbreeding schemes,
which allow an optimal use of maternal heterosis in Fl sows, may also remain an
excellent solution in some situations where the efficiency of lean tissue growth
is not a priority such as the production of high eating quality meat (Gandemer
et al., 1992)or extensive systems under tropical environment (Delate et al.,
1991). Discontinuous crossbreeding schemes might also be interesting under
intensive management systems it the use of growth promotors such as porcine
somatotropin (pST) to growing pigs were allowed. It has been shown that the
effects of pST are much larger in fat genotypes such as MS, thus reducing their
handicap for production traits. In an experiment involving lean (Pietrain, PI),
fat (Meishan, MS), or intermediate genotypes (Pl
´
3/4 Large White
´
1/4 MS), referred to as crossbred (CR), females were injected between 60 and 100
kg live weight with 6 mg/d of pST and compared to control females receiving the
vehicle only (Bidanel et al., 1991b). Average daily gain increased similarly in
the three genotypes (125 g/d). Feed conversion ratio tended to decrease to a
higher extent in MS(-2.0 kg of feed/kg of gain) than in the other two genotypes
(-1.1 and - 0.9 kg of feed/ kg of gain for CR and PI, respectively). A
significant genotype x treatment interaction was also observed for backfat
thickness (BF) and fat, muscle, and bone development. Effects of pST in PI, CR,
and MS pigs were, respectively, W.2, -9.6 and -16.1 mm for BF and 3.0, 6.8 and
11.8% for carcass lean con lent. This result could enhance the use of Chinese
genes through discontinuous crossbreeding schemes (using highly prolific Fl sows
to produce 1/4 Chinese slaughter pigs). However, much uncertainly still remains
about the authorization of using pST, particularly within the EEC market.
IV-CON'CLUSION
In spite of an extremely narrow
genetic basis, which precludes any extrapolation to the breeds as t whole,
French Meishan and to a lesser extent Jiaxing have confirmed under intensive
husbandry conditions their exceptional reproductive ability and their very poor
lean growth potential. Significant results were abtained over the last 12 years
in France, especially in the domain of reproduction. However, much work remains
to be done before we are fully able to understand the particularlies of these
breeds. From this point of' view, the recent development of research programmes
on Chinese pigs in several countries and international cooperation on the
subject should lead to much more important progress over the next decade.
Chinese breeds could play an active
part in the improvement of the efficiency of the pig industry as suggested by
recent development of research and breeding projects in an increasing number of
countries. Among different possible strategies for taking advantage of prolific
Chinese breeds in various market conditions, the creation of composite lines
selected in favour of growth and body composition seems to be by far the most
promising. However, the improvement of pure Chinese breeds or the use of
discontinuous crossbreeding themes completed by pST administration should also
be taken into consideration.
V-REFERENCES
Bazer, F. W., Thatcher, W.W.,
Marlinal-Bott4, F. and Terqui, M. 1988.
J. Reprod. Fert., S3: 723-728
Bidancl, J. P. 1988.
Bases xoeechniques et gbaetiques de
1’utilisation en gevage intensil' des races prolifiques chinoiaes-cas du pore
Meishan. 194 p, Doctoral Thesis, Inalilut National Agronomique Paris – Grignon,
France
Bidanel, J. P. 1989. In: 21es
Journess Rech. Porcine en
France, Paris, 31 janvicr-2 fevrier 1989, Institut Technique du Porc, Paris, pp
361-366
Bidanel, J. P. 1990. Tn. 4th World
Congress on Genetics applied to Livestock Productinn, Edinburgh, UK, vol XV, pp
481-484
Bidanel, J. P. and Legault, C. 1986.
In: 37th Annual Meeting of the EAAP, Budapest, Hungary, September 1986,
Commission on pig production, 13p
Bidanel, J. P., Carilez, J. C. and
Legault, C. 1989. Genet. Sel. Evol. 21:S07-526
Bidanel, J. P., Caritez, J. C. and
Legault, C. 1990a. Pig News & In fornation 11:345 – 348
Bidanel, J. P., Caritez J. C. and
Legault, C. 1990b. In: Chinese pig symposium July 5-6, 1990, Touloase, France (Molenat
M, Legaelt C, eds), pp 67-82
Bidanel, J.P., Cari', J.C. and
Laganl, H. 1990c. In: Chinese pig symposium, July 5-6, 1990. Touloaae, France
(Molenat M, Legault C. eds), pp 109 – 110
Bidanel, J. P., Caritez, J. C. and
Legault, C.1990d. Genet. Sel. Evol. 22: 43l~445
Bidanel, J. P., Caritez, J. C. and
Legault, C. 199la. Pig News & Information 12: 239-243
Bidanel, J. P., Bonneau, M.,
Pointillart, A., Gruand, J., Mourot, J. and Demade, I 199lb. J. Anim. Sci, 69:
3511-3522
Bidanel, J. P., Caritez, J. C.,
Gruand, J. and Legault, C. 1992. Genet. Sel Evol: (in press)
Bolet, G., Martinat-Botte, F.,
Localelli, P., Gruand, J., Terqui, M. and Berthelot, F. 1986. Genet. Sel. Evol.
18: 333-342
Bonneau, M., Mourot, J., Noblet, J.,
Lefaucheur, L. and Bidaael, J. P. 1990.la: Chineae pig symposium, July 5-6,
1990, Toulouse, France (Molenat M, Legault C, eds), pp 199-213
Bousset, J., Legault, C. and Merat,
P. 1990. In: Chinese pig symposium, July 5-6, 1990, Toulouae, France (Molenat M,
Legault C, eds), pp 257-258.
Canope, I., Despois, E. and Mandonnet,
N. 1990. In: Chineae pig
symposium, July 5-6, 1990, Toulouse, France(Molenat M, Legault C, eds), pp
131-132.
Chappuis, J.P., Duval-lflah, Y
Ollivier, L. and Legault, C. 1984. Genet. Sel. Evol. 16: 385-390
Chowdhary, B. P Johansson, M.,
Chowdhary, M., Ellegren, H Gu, F., Andersson, L. and Gustavsson, I. 1991. In
situ hybridization mapping and RFLP analisys of the porcine alburmin (ALB) and
transferrin (TF) genes. Cytogenet. Cell.
Genet. (HGM 11).
Delate, J. J., Le Guyadec, D., Le
duot, P. and Duclos, J. M. 1991. In: 23es Journees Rech. Porcine en France,
Paris, 5-7 fevrier 1991. Institut Technique du Pore, Paris, pp 38 l-388
Depress, E., Caritez, J. C.,
Legault, C. and Lagant, H. 1990.
In: 22ea Journbes Rech. Porcine en
France, Paris, 30 janvier-ler fevrier 1990, Inatitut Technique du Porc, Paris,
pp 333-338
Despres, P., Martinat-Botte, F.,
Lagant, H., Terqui, M. and Legault, C. 1992. In: 24es Journees Rech. Porcine en
France, Paris, 4-6 fevrier 1990, Institut Technique du Porc, Paris, pp25-30
Dickerson, G. E. 1969. Anim. Breed.
Abstr. 37: 191-202.
Fevrier, C., Bourdon, D., Aumaitre,
A., Peiniau, J., Le Bretnn, Y., Jaguelin, Y., Meziere, N. and Blanchard, A.
1988. In: 4th international
Seminar on Digestive Physiology in Pigs, Jablonna, Poland 7-9 June, 1988.
Gandemer, G., Viau, M., Caritez, J.
C. and Legault, C. 1992, Meat Science 32: 105-1212
Gauthier, M. C., Legault, C. and
Caritez J. C. 1992. In: 24es
Journess Rech. Porcine en France, Paris, 4-6 fevrier 1990, Institut Technique du
Porc, Paris, pp337-344
Gueblez, R., Bruel, L. and Legault,
C. l987. In: 19es Journees de
la Recherche Porcine en France, Paris, 4-5fevrier 1987, Institut Technique du
Porc, Paris, pp25-32
Le Dividich, J., Mormede, P.,
Catheline, M. and Caritez., J. C. 1989. Biol. Neonat.
59: 268-277
Legault, C., Caritez, J. C., Gruand,
J. and Sellier, P. 1982. In: 14es Journees de la Recherche Porcine en France,
Paris, 3-4 fevrier 1982, Institut Technique du Porc, Paris, pp 127-135
Legault,
C. and Caritez, J. C. 1983. Genet. Sel. Evol. 15: 225-240
Legault, C., Sellier, P., Caritez,
J. C., Dando, P. and Gruand, J. 1985. Genet. Sel. Evol. 17:133-152
Looft, C., Krapoth, J. and Kalm, E.
)990. In: Chinese pig symposium, July 5-6, 1990, Toulouae, France (Molenat M,
Legault C, eds), pp 125-126.
Mandonnet, N., Le Roy, P., Caritez,
J. C., Elsen, J. M., Legault, C. and Bidanel, J.P. 1992.
In: 24es Journees Rech. Porcine en
France, Paris, 4-6 fevrier 1992, Institut Technique du Porc, Paris, pp 25-30
Muramatsu, S., Popescu, P. C., Otani,
T. and Tamada, S. 1990. In: Chinese pig symposium, July 5-6, 1990, Toulouse, France (Molenat M,
Legault C, eds), pp 105-106.
Meunier-Salaun, M. C., Gort, F.,
Prunier, A., Caritez, J. C. and Schouten, W. P. G. 1991. In: 23es Journees Rech.
Parcine en France, Paris, 5-7 fevrier 1991, Institut Tachnique du Porc, Paris,
pp 409-414
Mormede, P., Dantzer, R., Bluthe, R.
M, and Caritez, J. C. 1984. Genet. Se1. Evol. 16:85-102
Naveau, J. 1990. In: Chinese pig
symposium, July 5-6, 1990, Toulouse, France (Molenat M, Legault C, eds), pp
123-124.
Ollivier, L. and Renjifo J. 1991.
Genet. Sel Evol. 23: 235-248.
Ollivier, L., .Gellin, J., Popescu,
P. and Vaiman, M. 1992. In:
24es Journess Rech. Porcine en France, Paris, 4-6 fevrier 1992, Institut
Technique du Porc, Paris, pp 1-8
Poilvet, D., Bonneau,
M., Caritez, J. C. and Legault, C. 1990. In: Chinese pig symposium, July 5-6,
1990, Toulouse, France (Molenat M, Legault C, eds), pp237-238
Prunier, A., Caritez J. C. and
Bonneau, M. 1987. Ann.
Zootech. 36: 49-58
Rousset, S., Touraille, C., Solignat,
G. and Molenat, M. 1990. In:
Chinese pig symposium July 5-6, 1990, Toulouse, France (Molenat M, Legault C,
eds), pp 261-262.
Terqui, M., Bazer, F. W. and
Martinal- Botte, F. 1992. In: Chinese pig symposium, August 11-14, 1992, Harbin,
China
Touraille, C., Monin, G. and Legault,
C. 1989. Meat Science 25: 177-186
[回上頁 | 顯示此文件為可列印格式]
此文件提供者: shuYing - [評分 : 0.00 (0 票選) | 評分!]









