IN VITRO PRODUCTION OF RUMINANT EMBRYOS: RESULTS, LIMITS AND PERSPECTIVES
P. Mermillod*, Y. Locatelli, R. Dalbiès-Tran, S. Uzbekova, G. Baril, F. Guignot, C. Perreau, N. Poulin, J. L. Touzé, S. Pennetier, B. Schmaltz, and Y. Cognié
INRA, UMR Physiologie de la Reproduction et des Comportements, INRA, CNRS, Université de Tours, Haras Nationaux, 37380 Nouzilly, France
* Corresponding author e-mail : Pascal.Mermillod@tours.inra.fr
ABSTRACT
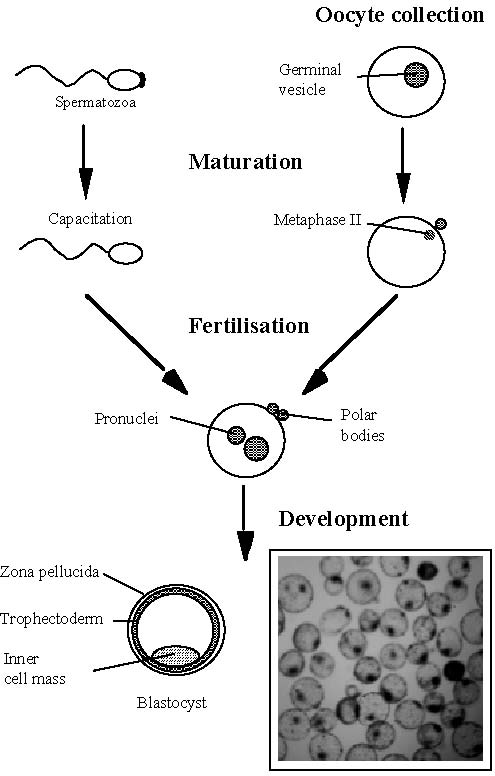
After artificial insemination and multiple ovulation and embryo transfer (MOET), in vitro production of embryos (IVP) represents the third generation of techniques aimed at a better control of animal reproduction. This technique involves four major steps (Figure 1): oocyte collection, oocyte in vitro maturation (IVM), in vitro fertilization (IVF) and in vitro development of the resulting embryos (IVD). These different steps are now well established in domestic ruminant species (cattle, sheep and goat) although the variability of the number and quality of the oocytes collected and the low viability of frozen – thawed in vitro produced embryos still limit the large-scale use of this promising technology. Beyond the potential use of IVP in breeding schemes, this technique is also required for the establishment of new biotechnologies such as cloning and animal transgenesis. Additionally, the knowledge of oocyte and embryo physiology acquired through IVP techniques may stimulate the further development of other techniques such as MOET.
KEY WORDS: Blastocyst, Development, Fertilization Freezing, Maturation, Oocyte, Transfer.
INTRODUCTION
The knowledge and use of reproductive technologies have been increasing worldwide all along the second part of the 19th century in most domestic mammalian species. First generation technologies (estrous synchronization, semen collection, semen freezing and artificial insemination) are now commonly used in domestic species breeding.
The second generation of reproductive techniques have then been introduced and their use have been promisingly increasing until early nineties. This generation included embryo based techniques, i.e. female superovulation, embryo recovery, freezing and transfer to recipients. At that time, as breeding industry was facing an increasing competition context worldwide, several health crisis, i.e. Bovine Spongiform Encephalopathy (BSE) and foot and mouth disease, negatively impacted the breeding economy and reduced the expansion of assisted reproductive techniques in European countries.
In addition, following these health crisis, there was an increasing rejection by consumers and public opinion of the use of animal originated products (such as pituitary extracts) in animal breeding and husbandry. This rational concern has been triggered by a more philosophical claim for “green” products obtained in highly natural conditions (i.e. with lowest as possible human intervention). This increasing demand for high quality and natural products by consumer, has progressively decreased the need for advanced reproductive techniques aimed at improving herds productivity.
Basically, IVP includes three major steps (Figure 1) : in vitro maturation (IVM), in vitro fertilization (IVF and in vitro development (IVD) of the resulting embryos. However, primary oocytes collection should be added upstream of these major steps and embryo management (freezing, transfer) should be added downstream to give a complete overview of the whole process. Since the birth of the first IVP calf in 1982, thanks to intensive research programs worldwide, cattle IVP has done significant progress. However, some residual shortcomings are still limiting the larger commercial use of this promising technique.
The focus of this review paper will be to describe the different steps of IVP and giving the results currently obtained. We will also describe the main limitations impairing a wider use of this promising technology and provide some research directions that may allow to overcome these limitations in the future. Bovine species will be presented as a general models but interesting particularities of other ruminant species will be also described.
OOCYTE COLLECTION
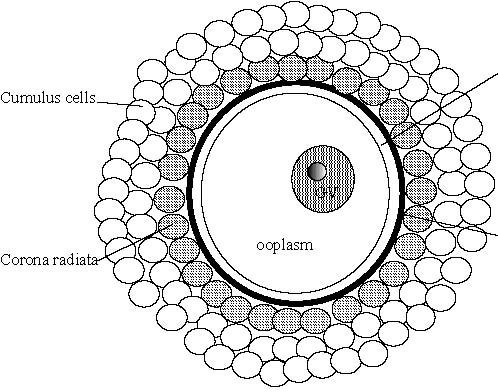
For the set up of the techniques as well as for the large scale production of average genetic merit embryos, large quantities of material can be obtained at low cost by collecting ovaries at slaughterhouses. These ovaries are shipped to the laboratory (dry or in warmed saline) where the content of the follicles is aspirated. The needle used as well as the aspiration vacuum are important factors in determining the number and quality of the oocytes collected. These parameters have to be established for each species. In the cow, good results can be obtained with 18 gauge needle connected to a 3 cm Hg vacuum. Alternatively, the ovaries may be sliced with a razor blade and washed with PBS to Corona radiatacollect the oocytes. An average of 5 to 10 oocytes can be collected from each bovine ovary (Mermillod et al. 1992) and one to two from sheep or goat ovaries during the breeding season (Cognie 1999). After aspiration or slicing, the collected fluid is screened under a stereomicroscope to select the oocytes. The oocytes are surrounded by several layers of somatic cells (cumulus oophorus, Figure 2). These cells establish contacts together as well as with the oocyte to allow cell communication through gap junctions, which is necessary for efficient maturation of the oocyte (Van Soom et al. 2002). Only cumulus oocyte complexes (COC) with complete and compact cumulus investment should be selected (Leibfried and First 1979; Stangl et al. 1999). Cumulus expansion observed at oocyte collection may reflect follicular atresia or premature meiotic resumption in the oocyte. Cresyl brillant blue vital staining could be used for better selection of COC according to the glucose-6-phosphate dehydrogenase activity in cumulus cells, that reflects their vitality and functionality (Pujol et al. 2004). The morphology of the ovaries themselves may also help to predict the developmental potential of enclosed oocytes (Gandolfi et al. 1997).
Figure 2. Schematic representation of a cumulus oocyte complex. Oocyte diameter is about 120 mm.
Figure 1. Major steps of in vitro embryo production (IVP)
To use this technique in a breeding scheme, the COC could be obtained from females of high genetic merit. This could be done after accidental culling of the female or at the end of her life to ensure a last reproduction (in the same way as described for slaughterhouse oocytes) or from living female, through the Ovum Pick Up (OPU) technique.
In the bovine, OPU is mainly done by transvaginal aspiration of follicle content under ultrasonographic control. This technique has been developed in humans and has been then adapted in cattle (Pieterse et al. 1988). Aspiration parameters are as important in OPU procedure than they are during aspiration of oocytes from slaughterhouse ovaries. Increasing aspiration flow increases oocyte recovery rate but decreases oocyte quality (Bols et al. 1997). The ratio of oocyte collected from aspirated follicles is usually around 60-70 %. OPU could be performed as frequently as twice a week, with or without hormonal stimulation of follicular growth, over several months without affecting further reproductive life of the donor (Bols, 2005; van Wagtendonk-de Leeuw, 2006). It has been shown that two OPU sessions per week allow the collection of oocytes of higher quality, translating into higher rate of development to the blastocyst stage as compared with one session per week (Chaubal et al. 2006). This may be due to the appearance between weekly sessions of a dominant follicle which negatively influences the quality of oocytes in subordinate follicles. Indeed, the removal of the dominant follicles by OPU between two weekly sessions increased the blastocyst yield. An average of ten oocytes per cow and 8 per heifer may be collected at each OPU session in a twice a week protocol without stimulation, resulting in an average of 2.5 transferable embryos per cow per session and 1.5 per heifer (Galli et al. 2003). This protocol presents the advantage to avoid the repeated use of exogenous gonadotropins which could induce several reproductive pathologies and may lead to subfertility. In goat, it has been shown that LOPU could be repeated up to five time at various intervals without decrease in efficiency (Pierson et al. 2005). However, we have shown that repetition of LOPU on a weekly basis in goat under pFSH stimulation progressively decreases oocyte recovery (Baril et al., unpublished data). This decrease may be due to the repeated starvation of the donors before LOPU.
Alternatively, OPU could be done on a weekly basis after FSH priming with similar yield of total good quality oocytes. After one shot FSH stimulation (80 mg i.m. and 120 mg s.c. of pFSH), it is possible to perform two successive OPU sessions at 30 h and 126 h after FSH injection with good embryo production rate (Chaubal et al. 2006). The combined intra muscular and subcutaneous administration of FSH allows a rapid stimulation (i.m.) as well as a sustained release of the hormone (s.c.) that avoid repeated handling of animals and ensure continuous follicular growth stimulation in a twice a week stimulation protocol. In the case of stimulated protocols with multiple injections (300 mg pFSH i.m., 6 injections 12 h apart), it has been shown that a coasting period of 48 h allows to increase the rate of oocytes reaching the blastocyst stage, specially when LH (Lutropin, Vetrepharm, Canada, i.v.) was administered 6 h before OPU (Blondin et al. 2002). We did not find similar effects of this coasting period in goat LOPU (Baril et al, unpublished results). In young heifers (2-3 months of age), good IVP results (up to 3 transferable embryos per calf) could be obtained after eCG stimulation followed by oocyte collection under laparotomy (Galli et al. 2004).
Table 1. Embryo production after OPU in a twice a week unstimulated protocol (adapted from Galli et al. 2003).
| OPUs | Oocyte (per OPU) | Cleavage | Embryos (per OPU) |
Cows | 2,641 | 26,532 (10.0) | 70.9 % | 7,403 (2.8) |
Heifers | 2,143 | 16,786 (7.8) | 67.0 % | 3,284 (1.5) |
Total | 4,784 | 43,318 | | 10,687 |
In small ruminants, OPU may be done by laparotomy or, less invasively, by laparoscopy guided aspiration of ovarian follicles (Baldassarre et al. 2002; Cognie et al. 2004). In laparoscopic ovum pick-up (LOPU), aspiration conditions (size of the needle, bevel shape, aspiration flow, tubing internal diameter) should also be controlled and adapted to the species, as in OPU, to improve recovery rate and oocyte quality (Rodriguez et al. 2006). Good results can be obtained by LOPU in goat and sheep with or without ovarian stimulation by FSH or eCG (Baldassarre et al. 2003; Morton et al. 2005).
In wild ruminant species, oocyte recovery technique should be adapted to the size of the donor. Transvaginal OPU could be used in large deer species such as wapiti and red deer (Berg and Asher, 2003), whereas LOPU is recommended for smaller animals like sika deer. In sika deer, it has been shown that LOPU after oFSH stimulation of ovarian activity allows the recovery of good quality oocytes and in vitro production of viable embryos all year long, even during the non breeding season (Locatelli et al. 2006).
IN VITRO MATURATION
Nuclear maturation
In the ovary, all primary oocytes are blocked at the prophase stage of the meiotic cycle The block of meiotic progression during follicular growth is controlled by unknown factors produced by the surrounding somatic cells (Mermillod and Marchal, 1999). At this stage, the round shaped nucleus, called the germinal vesicle (GV), contains diffuse chromatin. During early folliculogenesis, the oocyte grows, undergo strong morphological modifications and stores molecules (proteins, RNAs) that will be determinant for the success of fertilization and early embryo development. The zona pellucida, a glycoprotein envelope surrounding the oocyte membrane, is synthesized by the oocyte during this period of growth. The zona pellucida has important function in regulating fertilization and protection of the embryo until hatching. The oocytes resume meiosis and progress to the metaphase II stage only if they encounter a gonadotropin surge in the preovulatory follicle. They stop their progression at this MII stage until fertilization in the oviduct, after ovulation.
Oocytes that are collected from growing follicles for IVP are blocked at the prophase stage of meiosis As soon as they are removed from the follicular inhibitory environment, meiotic resumption occurs spontaneously and progress to metaphase II (Sirard, 2001). This spontaneous meiotic resumption of the oocyte outside of the follicle is the basis of in vitro maturation.
Cytoplasmic maturation
Cytoplasmic maturation covers all morphologic and molecular events accompanying nuclear maturation after LH surge in preovulatory follicles and preparing oocyte cytoplasm to successful fertilization and embryo development. Cytoplasmic maturation includes well known morphological modifications, such as the migration of cortical granules in the cortical region of the ooplasm (Cran, 1989). These granules are stored during oocyte growth and release their enzymatic content in the perivitelline space after fertilization. These enzymes modify the structure of the zona pellucida, preventing the penetration of additional spermatozoa. Meiotic competence is acquired during early folliculogenesis, soon after the apparition of the antral cavity in the follicle (Mermillod et al. 1999). However, after this acquisition, the oocyte requires a further differentiation period during late follicular growth to reach the full competence for cytoplasmic maturation (Sirard et al. 2006). This late differentiation occurs under the meiosis inhibiting signal sent by somatic follicular cells that maintain the meiotically competent oocyte at the prophase stage.
During IVP, more than 90% of the oocytes collected from follicles larger than 3 mm in cattle are able to complete nuclear maturation. However, few of them are competent for cytoplasmic maturation. Consequently, only 30 to 40% of the oocytes reach the blastocyst stage after IVF and IVD (Figure 3). This contrast highlights the functional importance of cytoplasmic competence (Sirard et al. 2006). When oocytes harvested from larger follicles or in vivo matured oocytes are processed under the same IVF – IVD techniques, the success rate is increased (Hue et al. 1997; Lequarre et al. 2005; Marchal et al. 2002). Ovarian stimulation by gonadotropins increases the number of follicles growing to large size classes by protecting them against involution through the atresia process. Consequently, ovarian stimulation increases the number of competent oocytes collected through the OPU process.
Increasing developmental competence during late follicular growth is suspected to be related to storage in oocyte of transcripts that will be necessary for the success of early embryo development. Indeed, in ruminant species, embryo genome activity only starts after the third embryo cell cycle (8-16 Cell stage, (Memili et al. 1998). Transcription activity has been studied in growing oocytes by following ribosomal RNA synthesis (Hyttel et al. 2001). These studies allowed to establish that the oocyte is transcriptionally quiescent in primordial follicles. RNA synthesis appears in secondary follicles, peaks in tertiary follicles, at later stages of oocyte growth, and decreases thereafter to return to undetectable levels. Meiotic resumption in late preovulatory follicle is preceded by a short burst of RNA synthesis (Memili et al. 1998) which seems to be functionally important for the success or embryo development, as evidenced by -amanitin (inhibitor of RNA polymerase II) inhibition of transcription at this stage. Several maternal effect genes, such as Mater (Maternal Antigen That Embryo Require) or zygote arrest 1 (Zar1) have been identified in mouse oocytes (Tong et al. 2000; Wu et al. 2003). The storage of these maternal transcripts in oocytes is required to allow embryos to develop beyond the 2-Cell stage. Orthologues of these genes have been recently identified in oocytes from domestic species (Dalbies-Tran et al. 2005; Pennetier et al. 2006; Pennetier et al. 2004) and their expression profile has been characterized (Pennetier et al. 2006). In addition, cDNA arrays are used for a more comprehensive analysis of oocyte’s expression patterns. These studies rely on the use of both heterologous arrays (Dalbies-Tran and Mermillod, 2003) or specific arrays builded from SSH selected oocyte specific cDNA banks (Pennetier et al. 2005; Sirard et al. 2005; Vallee et al. 2006). These studies have allowed the identification of several hundreds of genes expressed in the bovine oocyte, some of them being preferentially expressed in oocyte (and sometimes testis) and not detected in other tissues. However, up to now, none of these genes as been identified as a key element of oocyte developmental competence acquisition during late folliculogenesis.
Figure 3. Success rates of the successive IVP steps.

In vitro maturation
In vitro maturation is probably the most critical part of the whole process of in vitro embryo production. Ruminant oocytes are usually matured at 39°C under a 5% CO2 in a humidified atmosphere. The optimal maturation time (more than 90% of the oocytes at the metaphase II stage) is 22-24h. Due to the high lipid content of oocytes cytoplasm, it is not possible to follow the progression of their nuclear status during culture. Additionally, the presence of the cumulus cells is required during IVM for efficient cytoplasmic maturation and these cells mask the oocyte. Consequently, the only visible sign of oocyte maturation during IVM is the expansion of the cumulus cells. These cells produce hyaluronic acid which is secreted and polymerized in the extracellular matrix, leading to the increase of intercellular space.
Several culture media have been proposed for IVM (MEM, Waymouth, Ham-F12). However, the most widely used seems to be the TCM199 medium, bicarbonate buffered and containing minerals, carbon and energy sources (glucose, glutamine) as well as vitamins and amino acids. The medium is generally supplemented with high molecular weight molecules that exert a surfactant effect (BSA) and provide hormones and growth factors (fetal calf serum, serum of female in estrus, follicular fluid). These complex additives from animal sources may raise sanitary questions due to possible presence of pathogen agents, and decrease the reproducibility of experiments. They may be successfully replaced by high molecular weight polymers (Ali and Sirard, 2002) for the surfactant effect (such as polyvinyl alcohol) and by cocktails of purified or recombinant hormones and growth factors (Lonergan et al. 1996).
Hormones are usually added to the maturation medium (FSH, LH, estradiol), in addition to biological fluids. Their exact function is not clearly established, althrough the effect of FSH on cumulus expansion and metabolism has been clearly established (Sutton-McDowall et al. 2004). Estradiol effect on oocyte is exerted through intracellular receptors and seem to be detrimental to meiotic progression, inducing chromosomal abnormalities (Beker-van Woudenberg et al. 2004). Growth hormone (GH) stimulates some aspects of cytoplasmic maturation (cortical granules migration) as well as nuclear maturation, resulting in a higher blastocyst yield (Bevers and Izadyar, 2002). GH action seems to be mediated by cumulus cells (Izadyar et al. 1997). Epidermal growth factor (EGF) has been shown to stimulate nuclear and cytoplasmic maturation in a wide variety of species including mouse, pig, cattle, deer goat and sheep (Abeydeera et al. 2000; Cognie et al. 2004; Comizzoli et al. 2001b; De La Fuente et al. 1999; Guler et al. 2000; Lonergan et al. 1996). High blastocysts yields could be obtained after maturation of cattle oocytes in TCM199 only supplemented by EGF. EGF action is exerted, at least partly, directly on the oocyte and recent studies showed that EGF family of growth factors (epinephrin, amphiregullin, betacellulin) could be produced by granulosa cells under gonadotropins stimulation and could thus mediate LH signaling inside the follicle, transmitting the ovulatory signal to the oocyte (Park et al. 2004). Insulin like growth factor-I (IGF-I) has also a positive effect on cytoplasmic maturation of cattle oocytes (Bevers and Izadyar 2002). Indeed, the regulation of IGF-I and II through the controlled expression and degradation of IGF binding proteins could be one of the major processes of control of late follicular growth (Mazerbourg et al. 2003).
Another important aspect of COC environment during IVM is the antioxydative potential of the IVM medium (Ali et al. 2003). As all living cells, oocyte is sensitive to reactive oxygen species (ROS) that could induce detrimental chemical reactions (lipid or protein peroxydation). To overcome possible ROS detrimental effects during culture, antioxydative molecules ( -mercaptoethanol, cystein, vitamin A C, E), divalent cations chelators (EDTA, taurine, hypotaurine, transferrine) or ROS scavengers (superoxyde dismutase, catalase) could be added to maturation medium (Blondin et al. 1997; Ikeda et al. 2005; Kobayashi et al. 2006). However, COC are capable to produce their own protection system by secreting ROS scavenger enzymes like superoxyde dismutase and catalase (Cetica et al. 2001) or through the production of intracellular reducers molecules like glutathion (de Matos and Furnus, 2000). Stimulating these endogeneous protection systems may help oocyte to survive challenging in vitro conditions. The addition into IVM medium of cysteamine, a cell permeable precursor of cysteine, itself involved in glutathion (GSH) synthesis, has been shown to increase GSH concentration in oocytes and, in turn, to increase the resulting developmental competence after IVM in different species (Cognie et al. 2004; de Matos and Furnus, 2000; de Matos et al. 2002; Gasparrini et al. 2003; Khatir and Anouassi, 2006). Surrounding cumulus cells are also contributing to establishment of high GSH content in the oocyte (Luciano et al. 2005). Therefore, stimulating cumulus cells may also help to protect oocytes against environment ROS.
It has been established for a long time that the presence of cumulus cells is required for the success of oocyte cytoplasmic maturation and fertilization (Gilchrist et al. 2004). Therefore, maintaining the viability and functionality of cumulus cells is likely to be another important challenge for IVM success. Oocyte secreted factors (ESF) are known to drive cumulus cell differentiation during follicular growth (Li et al. 2000) and to allow cumulus expansion during oocyte maturation (Dragovic et al. 2005). The precise identification of the active factors is not fully established. However, GDF-9 and BMP-15, two members of the TGF-family of growth factors, are specifically produced by the oocyte and their effect on cumulus cells, and in turn on matured oocyte developmental competence, have been recently established (Hussein et al. 2006). Since these factors are able to protect cumulus cells against apoptosis that appear in culture (Hussein et al. 2005), their addition to IVM medium may be recommended, at least to help at maintaining cumulus functionality in culture.
IN VITRO FERTILIZATION
The techniques used for in vitro fertilization are well established in cattle as well as in small ruminants. The media used for IVF are Tyrode derived saline supplemented with BSA, lactate and pyruvate and with a capacitating agent (Parrish et al. 1986). Sperm capacitation occurs physiologically in the female genital tract. This phenomenon involves modification of spermatozoa membrane as well as changes in its mobility properties that lead to spermatozoa fertilization ability. Many protocols support sperm capacitation in vitro. In cattle, heparin is widely used for this purpose, usually without pre treatment (Parrish et al. 1989). In small ruminant, spermatozoa are usually capacitated by preliminary treatment with serum collected from females in estrus (Cognie et al. 2004). Frozen semen is generally used and a selection step is required to enrich the sperm suspension in living and motile spermatozoa. In the swim-up method, spermatozoa are deposit at the bottom of a culture medium containing tube and allowed to swim in the medium. The top fraction, containing motile sperms is then collected after a given incubation time. Alternatively, motile sperms may be purified by centrifugation in a discontinuous density gradient (Percoll). The motile fraction is collected at the bottom of the tube after centrifugation.
The sperm concentration used for IVF ranges between 0.5 and 2x106 spermatozoa per ml in fertilization medium. This concentration should be optimized for each male and even for each ejaculate from a given male. The fertilization rate increases together with sperm concentration. However, after a given value, an increasing proportion of the oocytes become polyspermic. A compromise has to be established for high fertilization rate with low polyspermy rate. For better convenience, sperm and oocytes are usually co-incubated for 18-24h, although a six hours incubation time seems to be sufficient to reach the higher level of normal fertilization.
A strong male effect has been reported on the success rate of IVF and, in turn, IVF may be used as a tool to evaluate male (or ejaculate) fertility (Marquant-Le Guienne et al. 1990; Papadopoulos et al. 2005). In addition, heterologous oocytes may be used to evaluta ejaculate fertility of males from rare species (Comizzoli et al. 2001a). Different males may provide different cleavage rates. Additionally, males providing similar fertilization rates may also differ in their ability to promote the development to the blastocyst stage. These differences between males may be related to their respective ability to up regulate glucose metabolism during G1 phase at pronuclear stage (Comizzoli et al. 2003) and consequently to allow earlier onset of the S phase in the zygote, translating into earlier cleavage and higher development rate (Comizzoli et al. 2000a). Therefore, each male to be used in IVF has to be tested for optimized fertilization conditions and for its ability to promote embryo development. In the case of OPU-IVP with high genetic merit females, the bulls’ ejaculates to use should previously be tested on slaughterhouse oocytes.
IN VITRO DEVELOPMENT
In standard cell culture conditions, ruminant embryo development blocks at the 8-16 cell stage. This block was first overcome through the use of co-culture systems involving oviduct epithelial cells (Eyestone et al. 1991) or other cell lineage (Myers et al. 1994) in standard tissue culture media (TCM199). The effect of cells has not been clearly established. They may produce some growth factors positively regulating embryo development or remove toxic media components (Killian 2004). In a second step, new culture media have been designed to more exactly fit embryo requirements without the need for co-culture. One of the first media designed for this purpose was synthetic oviduct fluid or SOF (Fukui et al. 1991). The composition of SOF was established from the composition of bovine oviduct fluid. However, SOF or SOF modifications have also been successfully used in small ruminant species. More recently, sequential media have been proposed to fit the evolution of embryo requirements during early development (Gardner et al. 2000). For example, glucose seems to be toxic to early cleaving embryos, whereas morula need some glucose in the medium for further development (Donnay et al. 2002; Gardner et al. 2000).
Semi defined media represent an easy way to produce mammalian embryos in vitro. However, embryo quality (viability after cryopreservation and/or transfer to recipient) appears to be low after development in such systems (Garcia-Garcia et al. 2006). On the other hand, co-culture systems may be more complicated to manage but allow the production of higher quality embryos. Therefore, some commercial bovine IVP teams still use co-culture system or embryo development in the oviduct of transient recipient ewes. In this case, the embryos are transferred in the oviduct of synchronized ewes just after IVF and surgically collected seven days later (Galli et al. 2004). This IVP system allows the production of embryos with similar quality and cryotolerance as in vivo collected ones. Recently, as we were trying to produce deer embryos, we observed that these embryos were not able to develop beyond the morula stage in the conventional SOF development system we use in our laboratory for different domestic ruminant species. To allow sika and red deer embryos to reach the blastocyst stage, we have been obliged to develop a co culture system involving ovine oviduct epithelial cell (oOEC) monolayers in SOF medium (Table 2). This culture systems allows the production of viable embryos since we obtained kids after transfer of fresh or frozen/thawed IVP red deer embryos produced in theses conditions (Locatelli et al. 2005; Locatelli et al. 2006). This observation in deer species highlight the function of the oviduct in the regulation of early embryo development and encourage research in view to identify and use oviduct originated embryotrophic factors to increase the efficiency of IVP (Killian, 2004).
Table 2. Cleavage on Day 2 and development on Day 8 of in vitro produced red and sika deer embryos in SOF medium with or without ovine oviduct epithelial cell (oOEC) support (adapted from Locatelli et al. 2005; Locatelli et al. 2006).
Species | oOEC | Oocytes (n) | Cleavage (%) | Blastocysts (%) |
Red deer | - | 64 | 66 | 3 |
| + | 93 | 63 | 25 |
Sika deer | - | 62 | 84 | 0 |
| + | 68 | 82 | 28 |
Blastocyst rate was calculated from total oocyte entering into IVM.
The development usually takes place at 39°c in droplets of culture medium overlaid with oil for 7 to 9 days. A high density of embryos per volume unit (one embryo per μl) seems beneficial to development (Ferry et al. 1994). When embryos are cultured without cell support, the oxygen concentration in atmosphere should be reduced to 5% to avoid induction of oxidative stress on the embryos (Olson and Seidel, 2000; Voelkel and Hu, 1992). As during IVM, addition of antioxydant molecules may increase rate or quality of IVP embryos (Bing et al. 2003; Dalvit et al. 2005; Takahashi et al. 2002). Serum is usually added to the culture medium from the beginning or after the first cleavages (48h post insemination). The presence of serum accelerates the development of embryos but seems to negative effects on their quality (Rizos et al. 2003) translating into fetal and placental abnormalities (Large Offspring Syndrome, see below) reported during pregnancy (Thompson et al. 1995). The rate of cleavage is usually high (over 80% of the total oocytes), when appropriate bull and fertilization conditions are used. However, the rate of development to the blastocyst stage plateaus at around 40% of the total oocytes. It has been clearly established that oocyte quality (intrinsic quality of primary oocyte coupled to maturation conditions used) determine the rate of blastocyst production, whereas development environment is determinant for embryo viability (Rizos et al. 2002).
The pregnancy rates obtained after transfer of fresh IVP embryos are usually good (40 to 50% of calves born). Some problems of calf abnormality and perinatal mortality have been reported (Farin et al. 2004; Farin et al. 2006; Kruip et al. 2000). However, these problems are less frequent than those observed with cloned embryos and seem to be linked to in vitro environment and to the presence of serum in the culture medium that induces chromosomes aberrations, modifications of embryo gene expression pattern and metabolism (Lazzari et al. 2002; Lonergan et al. 2004; Rizos et al. 2003).
The main limitation to a wider use of OPU – IVP technique in breeding schemes is the low cryoresistance of the resulting embryos. Although well established techniques of embryo freezing are described in cattle, sheep and goat and provide good results of pregnancy rate after transfer of the thawed embryos, these techniques appeared sub-optimal for the cryopreservation of IVP embryos. Several alternative methods of embryo cryopreservation have been proposed to overcome the low viability of these embryos. Some vitrification procedures (high speed cooling with high concentration of cryoprotectants leading to amorphous solidification without ice crystals formation) appeared to partially solve the problem. However, the quality of the embryo which is determined by the culture condition, appears to be one of the critical factors. Similar pregnancy rates have been observed after 10% glycerol slow freezing for in vitro fertilized embryos grown in sheep oviduct than for in vivo derived embryos (Galli et al. 2004). This indicates that culture conditions still remain to be improved to better mimic the in vivo conditions.
POTENTIAL INTERESTS OF IVP
Mass embryo production
The mass production of embryos from oocytes collected in the ovaries of slaughtered females may be of interest in some breeding schemes. For example, ovaries from the same breed may be pooled at slaughterhouse and used to produce embryos of a defined breed without precise knowledge of their parental origin, except breed. These embryos produced at low cost may be used to produced valuable beef calves in dairy herds (“beef from dairy”). This allows to increase the financial value of calves in these herds and may help solving fertility problems in high producing dairy herds (Galli et al. 2003; Hansen, 2006). Similar methods could also be used to produce average genetic value low cost dairy or beef embryos to help the diffusion of genetic progress toward developing countries. A similar approach has been used with success to reconstitute genetics of herds culled for sanitary reasons due to infections like tuberculosis or to BSE suspicion (Galli et al. 2003). In addition, IVP from slaughterhouse oocytes represent the most convenient way to develop basic research on oocyte maturation and cell cycle regulation, gametes recognition and fusion and regulation of early embryo development. Most recent knowledge of gene expression patterns and gene function in these mechanisms have been made possible thanks to slaughterhouse material.
Production of embryos from selected donors
Oocyte collection by OPU coupled to IVP offers several advantages over more traditional in vivo embryo production after superovulation and artificial insemination as applied in MOET schemes (Bols, 2005; Galli et al. 2004). In 2003, the International Embryo Transfer Society (IETS, reported 106,220 transfers of IVP cattle embryos worldwide (Hansen, 2006). Indeed, the number of embryos produced through OPU-IVP technique from a given female over a given period of time is higher than the number that may be collected in vivo. Indeed, OPU could be performed twice a week for long period of time without altering the reproductive potential of the females whereas several weeks or months of rest are required between two treatments of ovarian stimulation. In addition, in small ruminants, the surgery required for embryo collection in vivo induces tissue adhesions that could impair future embryo production, whereas LOPU is less invasive and could be repeated during long periods of time (Baldassarre and Karatzas, 2004). OPU can be performed in young females (from 3 months in age through laparoscopy) as well as in old cows. In addition, OPU could be continued during the first trimester of pregnancy without risk for the fetus, and it could be also performed on animals with some pathologies of the ovary (ovarian cysts), oviduct or uterus that prevent in vivo embryo production or even compromise fertility (Galli et al. 2003). OPU-IVP also allows the use of different males for producing the embryos (different males could be used for the oocytes from one aspiration session or from different OPU sessions). Furthermore, OPU-IVP could be beneficial in the case of females which are not responding to ovarian stimulation.
Wild species
Residual population of wild species maintained in captivity often encounter reproductive disorders that impair the amplification of the herd in view of reintroduction of animals in their natural environment or for the simple maintenance of these captive residual populations (Comizzoli et al. 2000b; Pukazhenthi et al. 2006). In addition, techniques of assisted reproduction like artificial insemination or embryo transfer may help to maintain proper management of scarce genetic diversity of few remaining individuals scattered between different locations. In this view, LOPU-IVP presents several advantages over MOET, like in domestic species and, in addition, it allows to reduce stress generating manipulations of wild animals (Locatelli et al. 2006).
MAJOR LIMITATIONS AND PERSPECTIVES
There are still some limitations to the extended commercial use of IVP (Faber et al. 2003; Hansen, 2006). The low success rate of development to the blastocyst stage is increasing the cost of each embryo produced. Additionally, some females appear to be poor oocyte donors in terms on both quantity and quality (developmental potential). The embryos produced by this technique have a low viability following cryopreservation and should be transferred fresh to get an optimized pregnancy rate. This limitation implies the disposition of a large recipient herds and the synchronization of recipients at each oocyte collection to prepare fresh transfer of resulting embryos, and consequently increases the overall cost of the technique. In addition, the possibility of embryos transportation is limited and embryo storage is not recommended.
Active research in several directions may help to overcome these limitations in a near future. Manipulations of ovarian stimulation protocols have shown that increasing late intrafollicular differentiation of the oocyte allows to increase its developmental potential. For example, introducing a coasting period after FSH stimulation of ovarian activity increases the rate of oocyte development to the blastocyst stage up to 80% (Blondin et al. 2002). This late differentiation appears to be critical to oocyte competence (Lequarre et al. 2005) and several authors tried to mimic this differentiation in vitro. Since meiotic resumption appears spontaneously in vitro, after suppression of follicle induced meiotic inhibition, and rapidly blocks transcriptions suspected to support developmental competence, mimicking late oocyte differentiation in vitro requires artificial maintenance of meiotic block (Mermillod et al. 1999). Attempts to recreate in vitro a follicular-like environment by adding follicular cells together with COC, provided limited successes (Sirard, 2001; Sirard and Bilodeau, 1990). More recently, it has been established that kinase inhibitors specifically blocking the kinase activity of M-phase Promoting Factor (MPF, key regulator of G2 – M transition of the eukayotic cell cycle) were able to reversibly maintain cattle oocyte at the GV stage in culture (Lonergan et al. 2000; Mermillod et al. 2000). However, these techniques of meiotic inhibition failed to increase l competence after a 24 h pre maturation culture of the oocytes at the GV stage. Nevertheless, these approaches allowed to establish that numerous signaling pathways are involved in oocyte cell cycle control (Vigneron et al. 2003; Vigneron et al. 2004a; Vigneron et al. 2004b). The increasing knowledge of this control will allow to derive more valuable methods of oocyte meiotic inhibition in the future. In addition, future studies will benefit from the increasing knowledge of the expression of genes related to developmental competence of the oocyte (Albertini et al. 2003).
Finally, some abnormalities have been reported during pregnancies after transfer of IVP embryos that have been called large offspring syndrome or LOS (Farin et al. 2006). This translates into abnormally large offspring after longer gestation time and higher rate of perinatal mortality. In sheep, this syndrome has been clearly related to the presence of serum in embryo culture medium (van Wagtendonk-de Leeuw et al. 2000) and may result from the maintenance of gene expression alterations induced by culture environment during early development stages (Lazzari et al. 2002). These problems may be overcome by using more physiological conditions for IVD. This may be achieved by developing the embryos into the oviduct of transient recipient ewes (Galli et al. 2004). However, recent progress in the comprehension of oviduct physiology (Killian, 2004) may allow to develop in vitro culture conditions mimicking in vivo environment to increase embryo quality. In addition, the increasing knowledge of embryo gene expression pattern and its regulation will allow to use marker genes to follow more precisely the effect of modification of embryo environment (Lonergan et al. 2006).
CONCLUSIONS
In conclusion, IVP is a very promising technique that could find numerous applications in assisted reproduction of ruminant domestic and wild species. It also offers unique occasion to study early mechanisms controlling reproductive physiology and open the way to advanced biotechnology such as nuclear transfer cloning and transgenesis. The low quality of oocytes recovered from small growing follicles as well as the low resistance of IVP embryos to conventional freezing methods still limit the use of IVP in breeding schemes. Improving IVP conditions in the future will require strong research efforts in view to improve our knowledge of oocyte and early embryo physiology and gene expression and to use this knowledge to develop in vitro treatments more adapted to precise requirements of oocytes and early embryos.
REFERENCE
Abeydeera L. R, Wang W. H, Cantley T. C, Rieke A, Murphy CN, Prather RS, Day BN. 2000. Development and viability of pig oocytes matured in a protein-free medium containing epidermal growth factor. Theriogenology 54(5):787-797.
Albertini D. F, Sanfins A, Combelles C. M. 2003. Origins and manifestations of oocyte maturation competencies. Reprod Biomed Online 6(4):410-415.
Ali A, Sirard M. A. 2002. Effect of the absence or presence of various protein supplements on further development of bovine oocytes during in vitro maturation. Biol Reprod 66(4):901-905.
Ali A. A, Bilodeau J. F, Sirard M. A. 2003. Antioxidant requirements for bovine oocytes varies during in vitro maturation, fertilization and development. Theriogenology 59(3-4):939-949.
Baldassarre H, Karatzas C. N. 2004. Advanced assisted reproduction technologies (ART) in goats. Anim Reprod Sci 82-83:255-266.
Baldassarre H, Keefer C, Wang B, Lazaris A, Karatzas C. N. 2003. Nuclear transfer in goats using in vitro matured oocytes recovered by laparoscopic ovum pick-up. Cloning Stem Cells 5(4):279-285.
Baldassarre H, Wang B, Kafidi N, Keefer C, Lazaris A, Karatzas C. N. 2002. Advances in the production and propagation of transgenic goats using laparoscopic ovum pick-up and in vitro embryo production technologies. Theriogenology 57(1):275-284.
Beker-van Woudenberg A. R, van Tol H. T, Roelen B. A, Colenbrander B, Bevers M. M. 2004. Estradiol and its membrane-impermeable conjugate (estradiol-bovine serum albumin) during in vitro maturation of bovine oocytes: effects on nuclear and cytoplasmic maturation, cytoskeleton, and embryo quality. Biol Reprod 70(5):1465-1474.
Berg D. K, Asher G. W. 2003. New developments reproductive technologies in deer. Theriogenology 59(1):189-205.
Bevers M. M, Izadyar F. 2002. Role of growth hormone and growth hormone receptor in oocyte maturation. Mol Cell Endocrinol 197(1-2):173-178.
Bing Y. Z, Hirao Y, Takenouchi N, Che L. M, Nakamura H, Yodoi J, Nagai T. 2003. Effects of thioredoxin on the preimplantation development of bovine embryos. Theriogenology 59(3-4):863-873.
Blondin P, Bousquet D, Twagiramungu H, Barnes F, Sirard M. A. 2002. Manipulation of follicular development to produce developmentally competent bovine oocytes. Biol Reprod 66(1):38-43.
Blondin P, Coenen K, Sirard M. A. 1997. The impact of reactive oxygen species on bovine sperm fertilizing ability and oocyte maturation. J Androl 18(4):454-460.
Bols P. E. 2005. Puncture of immature ovarian follicles in bovine assisted reproduction. Verh K Acad Geneeskd Belg 67(3):177-202.
Bols P. E, Ysebaert M. T, Van Soom A, de Kruif A. 1997. Effects of needle tip bevel and aspiration procedure on the morphology and developmental capacity of bovine compact cumulus oocyte complexes. Theriogenology 47(6):1221-1236.
Cetica P. D, Pintos L. N, Dalvit G. C, Beconi M. T. 2001. Antioxidant enzyme activity and oxidative stress in bovine oocyte in vitro maturation. IUBMB Life 51(1):57-64.
Chaubal S. A, Molina J. A, Ohlrichs C. L, Ferre L. B, Faber D. C, Bols P. E, Riesen J. W, Tian X, Yang X. 2006. Comparison of different transvaginal ovum pick-up protocols to optimise oocyte retrieval and embryo production over a 10-week period in cows. Theriogenology 65(8):1631-1648.
Cognie Y. 1999. State of the art in sheep-goat embryo transfer. Theriogenology 51(1):105-116.
Cognie Y, Poulin N, Locatelli Y, Mermillod P. 2004. State-of-the-art production, conservation and transfer of in-vitro-produced embryos in small ruminants. Reprod Fertil Dev 16(4):437-445.
Comizzoli P, Marquant-Le Guienne B, Heyman Y, Renard J. P. 2000a. Onset of the first S-phase is determined by a paternal effect during the G1-phase in bovine zygotes. Biol Reprod 62(6):1677-1684.
Comizzoli P, Mauget R, Mermillod P. 2001a. Assessment of in vitro fertility of deer spermatozoa by heterologous IVF with zona-free bovine oocytes. Theriogenology 56(2):261-274.
Comizzoli P, Mermillod P, Cognie Y, Chai N, Legendre X, Mauge R. 2001b. Successful in vitro production of embryos in the red deer (Cervus elaphus) and the sika deer (Cervus nippon). Theriogenology 55(2):649-659.
Comizzoli P, Mermillod P, Mauget R. 2000b. Reproductive biotechnologies for endangered mammalian species. Reprod Nutr Dev 40(5):493-504.
Comizzoli P, Urner F, Sakkas D, Renard J. P. 2003. Up-regulation of glucose metabolism during male pronucleus formation determines the early onset of the s phase in bovine zygotes. Biol Reprod 68(5):1934-1940.
Cran D. G. 1989. Cortical granules during oocyte maturation and fertilization. J Reprod Fertil Suppl 38:49-62.
Dalbies-Tran R, Mermillod P. 2003. Use of heterologous complementary DNA array screening to analyze bovine oocyte transcriptome and its evolution during in vitro maturation. Biol Reprod 68(1):252-261.
Dalbies-Tran R, Papillier P, Pennetier S, Uzbekova S, Monget P. 2005. Bovine mater-like NALP9 is an oocyte marker gene. Mol Reprod Dev 71(4):414-421.
Dalvit G. C, Cetica P. D, Pintos L. N, Beconi M. T. 2005. Reactive oxygen species in bovine embryo in vitro production. Biocell 29(2):209-212.
De La Fuente R, O'Brien M. J, Eppig J. J. 1999. Epidermal growth factor enhances preimplantation developmental competence of maturing mouse oocytes. Hum Reprod 14(12):3060-3068.
de Matos D. G, Furnus C. C. 2000. The importance of having high glutathione (GSH) level after bovine in vitro maturation on embryo development effect of beta-mercaptoethanol, cysteine and cystine. Theriogenology 53(3):761-771.
de Matos D. G, Gasparrini B, Pasqualini S. R, Thompson J. G. 2002. Effect of glutathione synthesis stimulation during in vitro maturation of ovine oocytes on embryo development and intracellular peroxide content. Theriogenology 57(5):1443-1451.
Donnay I, Feugang J. M, Bernard S, Marchandise J, Pampfer S, Moens A, Dessy F. 2002. Impact of adding 5.5 mM glucose to SOF medium on the development, metabolism and quality of in vitro produced bovine embryos from the morula to the blastocyst stage. Zygote 10(3):189-199.
Dragovic R. A, Ritter L. J, Schulz S. J, Amato F, Armstrong D. T, Gilchrist R. B. 2005. Role of oocyte-secreted growth differentiation factor 9 in the regulation of mouse cumulus expansion. Endocrinology 146(6):2798-2806.
Eyestone W. H, Jones J. M, First N. L. 1991. Some factors affecting the efficacy of oviduct tissue-conditioned medium for the culture of early bovine embryos. J Reprod Fertil 92(1):59-64.
Faber D. C, Molina J. A, Ohlrichs C. L, Vander Zwaag D. F, Ferre L. B. 2003. Commercialization of animal biotechnology. Theriogenology 59(1):125-138.
Farin C. E, Farin P. W, Piedrahita J. A. 2004. Development of fetuses from in vitro-produced and cloned bovine embryos. J Anim Sci 82 E-Suppl:E53-62.
Farin P. W, Piedrahita J. A, Farin C. E. 2006. Errors in development of fetuses and placentas from in vitro-produced bovine embryos. Theriogenology 65(1):178-191.
Ferry L, Mermillod P, Massip A, Dessy F. 1994. Bovine embryos cultured in serum-poor oviduct-conditioned medium need cooperation to reach the blastocyst stage. Theriogenology 42(3):445-453.
Fukui Y, McGowan L. T, James R. W, Pugh P. A, Tervit H. R. 1991. Factors affecting the in-vitro development to blastocysts of bovine oocytes matured and fertilized in vitro. J Reprod Fertil 92(1):125-131.
Galli C, Duchi R, Crotti G, Turini P, Ponderato N, Colleoni S, Lagutina I, Lazzari G. 2003. Bovine embryo technologies. Theriogenology 59(2):599-616.
Galli C, Duchi R, Crotti G, Turini P, Ponderato N, Colleoni S, Lagutina I, Lazzari G. 2004. Production and quality of bovine oocytes and embryos. Vet Res Commun 28 Suppl 1:121-126.
Gandolfi F, Luciano A. M, Modina S, Ponzini A, Pocar P, Armstrong D. T, Lauria A. 1997. The in vitro developmental competence of bovine oocytes can be related to the morphology of the ovary. Theriogenology 48(7):1153-1160.
Garcia-Garcia R. M, Ward F, Fair S, O'Meara C M, Wade M, Duffy P, Lonergan P. 2006. Development and quality of sheep embryos cultured in commercial G1.3/G2.3 sequential media. Anim Reprod Sci.
Gardner D. K, Pool T. B, Lane M. 2000. Embryo nutrition and energy metabolism and its relationship to embryo growth, differentiation, and viability. Semin Reprod Med 18(2):205-218.
Gasparrini B, Sayoud H, Neglia G, Matos D. G, Donnay I, Zicarelli L. 2003. Glutathione synthesis during in vitro maturation of buffalo (Bubalus bubalis) oocytes: effects of cysteamine on embryo development. Theriogenology 60(5):943-952.
Gilchrist R. B, Ritter L. J, Armstrong D. T. 2004. Oocyte-somatic cell interactions during follicle development in mammals. Anim Reprod Sci 82-83:431-446.
Guler A, Poulin N, Mermillod P, Terqui M, Cognie Y. 2000. Effect of growth factors, EGF and IGF-I, and estradiol on in vitro maturation of sheep oocytes. Theriogenology 54(2):209-218.
Hansen P. J. 2006. Realizing the promise of IVF in cattle--an overview. Theriogenology 65(1):119-125.
Hue I, Dedieu T, Huneau D, Ruffini S, Gall L, Crozet N. 1997. Cyclin B1 expression in meiotically competent and incompetent goat oocytes. Mol Reprod Dev 47(2):222-228.
Hussein T. S, Froiland D. A, Amato F, Thompson J. G, Gilchrist R. B. 2005. Oocytes prevent cumulus cell apoptosis by maintaining a morphogenic paracrine gradient of bone morphogenetic proteins. J Cell Sci 118(Pt 22):5257-5268.
Hussein T. S, Thompson J. G, Gilchrist R. B. 2006. Oocyte-secreted factors enhance oocyte developmental competence. Dev Biol.
Hyttel P, Viuff D, Fair T, Laurincik J, Thomsen P. D, Callesen H, Vos P. L, Hendriksen P. J, Dieleman SJ, Schellander K, Besenfelder U, Greve T. 2001. Ribosomal RNA gene expression and chromosome aberrations in bovine oocytes and preimplantation embryos. Reproduction 122(1):21-30.
Ikeda S, Kitagawa M, Imai H, Yamada M. 2005. The roles of vitamin A for cytoplasmic maturation of bovine oocytes. J Reprod Dev 51(1):23-35.
Izadyar F, Van Tol H. T, Colenbrander B, Bevers M. M. 1997. Stimulatory effect of growth hormone on in vitro maturation of bovine oocytes is exerted through cumulus cells and not mediated by IGF-I. Mol Reprod Dev 47(2):175-180.
Khatir H, Anouassi A. 2006. The first dromedary (Camelus dromedarius) offspring obtained from in vitro matured, in vitro fertilized and in vitro cultured abattoir-derived oocytes. Theriogenology 65(9):1727-1736.
Killian G. J. 2004. Evidence for the role of oviduct secretions in sperm function, fertilization and embryo development. Anim Reprod Sci 82-83:141-153.
Kobayashi M, Lee E. S, Fukui Y. 2006. Cysteamine or beta-mercaptoethanol added to a defined maturation medium improves blastocyst formation of porcine oocytes after intracytoplasmic sperm injection. Theriogenology 65(6):1191-1199.
Kruip T. A, Bevers M. M, Kemp B. 2000. Environment of oocyte and embryo determines health of IVP offspring. Theriogenology 53(2):611-618.
Lazzari G, Wrenzycki C, Herrmann D, Duchi R, Kruip T, Niemann H, Galli C. 2002. Cellular and molecular deviations in bovine in vitro-produced embryos are related to the large offspring syndrome. Biol Reprod 67(3):767-775.
Leibfried L, First N. L. 1979. Characterization of bovine follicular oocytes and their ability to mature in vitro. J Anim Sci 48(1):76-86.
Lequarre A. S, Vigneron C, Ribaucour F, Holm P, Donnay I, Dalbies-Tran R, Callesen H, Mermillod P. 2005. Influence of antral follicle size on oocyte characteristics and embryo development in the bovine. Theriogenology 63(3):841-859.
Li R, Norman R. J, Armstrong D. T, Gilchrist R. B. 2000. Oocyte-secreted factor(s) determine functional differences between bovine mural granulosa cells and cumulus cells. Biol Reprod 63(3):839-845.
Locatelli Y, Cognie Y, Vallet J. C, Baril G, Verdier M, Poulin N, Legendre X, Mermillod P. 2005. Successful use of oviduct epithelial cell coculture for in vitro production of viable red deer (Cervus elaphus) embryos. Theriogenology 64(8):1729-1739.
Locatelli Y, Vallet J. C, Huyghe F. P, Cognie Y, Legendre X, Mermillod P. 2006. Laparoscopic ovum pick-up and in vitro production of sika deer embryos: Effect of season and culture conditions. Theriogenology.
Lonergan P, Carolan C, Van Langendonckt A, Donnay I, Khatir H, Mermillod P. 1996. Role of epidermal growth factor in bovine oocyte maturation and preimplantation embryo development in vitro. Biol Reprod 54(6):1420-1429.
Lonergan P, Dinnyes A, Fair T, Yang X, Boland M. 2000. Bovine oocyte and embryo development following meiotic inhibition with butyrolactone I. Mol Reprod Dev 57(2):204-209.
Lonergan P, Fair T, Corcoran D, Evans A. C. 2006. Effect of culture environment on gene expression and developmental characteristics in IVF-derived embryos. Theriogenology 65(1):137-152.
Lonergan P, Pedersen H. G, Rizos D, Greve T, Thomsen P. D, Fair T, Evans A, Boland M. P. 2004. Effect of the post-fertilization culture environment on the incidence of chromosome aberrations in bovine blastocysts. Biol Reprod 71(4):1096-1100.
Luciano A. M, Lodde V, Beretta M. S, Colleoni S, Lauria A, Modina S. 2005. Developmental capability of denuded bovine oocyte in a co-culture system with intact cumulus-oocyte complexes: role of cumulus cells, cyclic adenosine 3',5'-monophosphate, and glutathione. Mol Reprod Dev 71(3):389-397.
Marchal R, Vigneron C, Perreau C, Bali-Papp A, Mermillod P. 2002. Effect of follicular size on meiotic and developmental competence of porcine oocytes. Theriogenology 57(5):1523-1532.
Marquant-Le Guienne B, Humblot P, Thibier M, Thibault C. 1990. Evaluation of bull semen fertility by homologous in vitro fertilization tests. Reprod Nutr Dev 30(2):259-266.
Mazerbourg S, Bondy C. A, Zhou J, Monget P. 2003. The insulin-like growth factor system: a key determinant role in the growth and selection of ovarian follicles? a comparative species study. Reprod Domest Anim 38(4):247-258.
Memili E, Dominko T, First N. L. 1998. Onset of transcription in bovine oocytes and preimplantation embryos. Mol Reprod Dev 51(1):36-41.
Mermillod P, Marchal R. 1999. [Oocyte of domestic mammals: a model for the study of in vitro maturation]. Contracept Fertil Sex 27(6):440-448.
Mermillod P, Oussaid B, Cognie Y. 1999. Aspects of follicular and oocyte maturation that affect the developmental potential of embryos. J Reprod Fertil Suppl 54:449-460.
Mermillod P, Tomanek M, Marchal R, Meijer L. 2000. High developmental competence of cattle oocytes maintained at the germinal vesicle stage for 24 hours in culture by specific inhibition of MPF kinase activity. Mol Reprod Dev 55(1):89-95.
Mermillod P, Wils C, Massip A, Dessy F. 1992. Collection of oocytes and production of blastocysts in vitro from individual, slaughtered cows. J Reprod Fertil 96(2):717-723.
Morton K M, de Graaf S. P, Campbell A, Tomkins L. M, Maxwell W. M, Evans G. 2005. Repeat ovum pick-up and in vitro embryo production from adult ewes with and without FSH treatment. Reprod Domest Anim 40(5):422-428.
Myers M. W, Broussard J. R, Menezo Y, Prough S. G, Blackwell J, Godke R. A, Thibodeaux J. K. 1994. Established cell lines and their conditioned media support bovine embryo development during in-vitro culture. Hum Reprod 9(10):1927-1931.
Olson S. E, Seidel G. E, Jr. 2000. Reduced oxygen tension and EDTA improve bovine zygote development in a chemically defined medium. J Anim Sci 78(1):152-157.
Papadopoulos S, Hanrahan J. P, Donovan A, Duffy P, Boland M. P, Lonergan P. 2005. In vitro fertilization as a predictor of fertility from cervical insemination of sheep. Theriogenology 63(1):150-159.
Park J. Y, Su YQ, Ariga M, Law E, Jin S. L, Conti M. 2004. EGF-like growth factors as mediators of LH action in the ovulatory follicle. Science 303(5658):682-684.
Parrish J. J, Susko-Parrish J. L, First N. L. 1989. Capacitation of bovine sperm by heparin: inhibitory effect of glucose and role of intracellular pH. Biol Reprod 41(4):683-699.
Parrish J. J, Susko-Parrish J. L, Leibfried-Rutledge M. L, Critser E. S, Eyestone W. H, First N. L. 1986. Bovine in vitro fertilization with frozen-thawed semen. Theriogenology 25(4):591-600.
Pennetier S, Perreau C, Uzbekova S, Thelie A, Delaleu B, Mermillod P, Dalbies-Tran R. 2006. MATER protein expression and intracellular localization throughout folliculogenesis and preimplantation embryo development in the bovine. BMC Dev Biol 6:26.
Pennetier S, Uzbekova S, Guyader-Joly C, Humblot P, Mermillod P, Dalbies-Tran R. 2005. Genes preferentially expressed in bovine oocytes revealed by subtractive and suppressive hybridization. Biol Reprod 73(4):713-720.
Pennetier S, Uzbekova S, Perreau C, Papillier P, Mermillod P, Dalbies-Tran R. 2004. Spatio-temporal expression of the germ cell marker genes MATER, ZAR1, GDF9, BMP15,andVASA in adult bovine tissues, oocytes, and preimplantation embryos. Biol Reprod 71(4):1359-1366.
Pierson J, Wang B, Neveu N, Sneek L, Cote F, Karatzas C. N, Baldassarre H. 2005. Effects of repetition, interval between treatments and season on the results from laparoscopic ovum pick-up in goats. Reprod Fertil Dev 16(8):795-799.
Pieterse M. C, Kappen K. A, Kruip T. A, Taverne M. A. 1988. Aspiration of bovine oocytes during transvaginal ultrasound scanning of the ovaries. Theriogenology 30(4):751-762.
Pujol M, Lopez-Bejar M, Paramio M. T. 2004. Developmental competence of heifer oocytes selected using the brilliant cresyl blue (BCB) test. Theriogenology 61(4):735-744.
Pukazhenthi B, Comizzoli P, Travis A. J, Wildt D. E. 2006. Applications of emerging technologies to the study and conservation of threatened and endangered species. Reprod Fertil Dev 18(1-2):77-90.
Rizos D, Gutierrez-Adan A, Perez-Garnelo S, De La Fuente J, Boland M. P, Lonergan P. 2003. Bovine embryo culture in the presence or absence of serum: implications for blastocyst development, cryotolerance, and messenger RNA expression. Biol Reprod 68(1):236-243.
Rizos D, Ward F, Duffy P, Boland M. P, Lonergan P. 2002. Consequences of bovine oocyte maturation, fertilization or early embryo development in vitro versus in vivo: implications for blastocyst yield and blastocyst quality. Mol Reprod Dev 61(2):234-248.
Rodriguez C, Anel L, Alvarez M, Anel E, Boixo J. C, Chamorro C. A, de Paz P. 2006. Ovum pick-up in sheep: a comparison between different aspiration devices for optimal oocyte retrieval. Reprod Domest Anim 41(2):106-113.
Sirard M. A. 2001. Resumption of meiosis: mechanism involved in meiotic progression and its relation with developmental competence. Theriogenology 55(6):1241-1254.
Sirard M. A, Bilodeau S. 1990. Granulosa cells inhibit the resumption of meiosis in bovine oocytes in vitro. Biol Reprod 43(5):777-783.
Sirard M. A, Dufort I, Vallee M, Massicotte L, Gravel C, Reghenas H, Watson A. J, King W. A, Robert C. 2005. Potential and limitations of bovine-specific arrays for the analysis of mRNA levels in early development: preliminary analysis using a bovine embryonic array. Reprod Fertil Dev 17(1-2):47-57.
Sirard M. A, Richard F, Blondin P, Robert C. 2006. Contribution of the oocyte to embryo quality. Theriogenology 65(1):126-136.
Stangl M, Kuhholzer B, Besenfelder U, Brem G. 1999. Repeated endoscopic ovum pick-up in sheep. Theriogenology 52(4):709-716.
Sutton-McDowall M. L, Gilchrist R. B, Thompson J. G. 2004. Cumulus expansion and glucose utilisation by bovine cumulus-oocyte complexes during in vitro maturation: the influence of glucosamine and follicle-stimulating hormone. Reproduction 128(3):313-319.
Takahashi M, Nagai T, Okamura N, Takahashi H, Okano A. 2002. Promoting effect of beta-mercaptoethanol on in vitro development under oxidative stress and cystine uptake of bovine embryos. Biol Reprod 66(3):562-567.
Thompson J. G, Gardner D. K, Pugh P. A, McMillan W. H, Tervit H. R. 1995. Lamb birth weight is affected by culture system utilized during in vitro pre-elongation development of ovine embryos. Biol Reprod 53(6):1385-1391.
Tong Z. B, Gold L, Pfeifer K. E, Dorward H, Lee E, Bondy C. A, Dean J, Nelson L. M. 2000. Mater, a maternal effect gene required for early embryonic development in mice. Nat Genet 26(3):267-268.
Vallee M, Robert C, Methot S, Palin M. F, Sirard M. A. 2006. Cross-species hybridizations on a multi-species cDNA microarray to identify evolutionarily conserved genes expressed in oocytes. BMC Genomics 7:113.
Van Soom A, Tanghe S, De Pauw I, Maes D, de Kruif A. 2002. Function of the cumulus oophorus before and during mammalian fertilization. Reprod Domest Anim 37(3):144-151.
van Wagtendonk-de Leeuw A. M. 2006. Ovum pick up and in vitro production in the bovine after use in several generations: a 2005 status. Theriogenology 65(5):914-925.
van Wagtendonk-de Leeuw A. M, Mullaart E, de Roos A. P, Merton J. S, den Daas J. H, Kemp B, de Ruigh L. 2000. Effects of different reproduction techniques: AI MOET or IVP, on health and welfare of bovine offspring. Theriogenology 53(2):575-597.
Vigneron C, Nuttinck F, Perreau C, Reinaud P, Charpigny G, Mermillod P. 2003. Effect of roscovitine, a cdk1 inhibitor, and of the presence of oocyte on bovine cumulus cell expansion and cyclooxygenase-2 expression. Mol Reprod Dev 65(1):114-121.
Vigneron C, Perreau C, Dalbies-Tran R, Joly C, Humblot P, Uzbekova S, Mermillod P. 2004a. Protein synthesis and mRNA storage in cattle oocytes maintained under meiotic block by roscovitine inhibition of MPF activity. Mol Reprod Dev 69(4):457-465.
Vigneron C, Perreau C, Dupont J, Uzbekova S, Prigent C, Mermillod P. 2004b. Several signaling pathways are involved in the control of cattle oocyte maturation. Mol Reprod Dev 69(4):466-474.
Voelkel S. A, Hu Y. X. 1992. Effect of gas atmosphere on the development of one-cell bovine embryos in two culture systems. Theriogenology 37(5):1117-1131.
Wu X, Viveiros M. M, Eppig J. J, Bai Y, Fitzpatrick S. L, Matzuk M. M. 2003. Zygote arrest 1 (Zar1) is a novel maternal-effect gene critical for the oocyte-to-embryo transition. Nat Genet 33(2):187-191.








