|
|
[回上頁 | 顯示此文件為可列印格式]
此文件提供者: shuYing - [評分 : 0.00 (0 票選) | 評分!]
飼養專欄
調整非乳脂固形物的方法
臺灣大學動物科學技術學系 徐濟泰 教授
先說一則故事。很多年前在某個場合聽到一位前輩的演講,他說國際貿易很像國際體育賽事,我們身高比較矮無法與西方國家競爭籃球,而棒球、舉重等比較不受身高影響,所以我們要培養的是棒球、舉重等參加國際比賽好拿個好成績。這樣的比喻淺顯易懂,三言兩語說明了國際現實,卻也道盡了我們的辛酸:如果天生條件不如人,你就不要打球了。世界不僅是平的,近年又流行區域整合追求最高效益,農業本來是兼顧生產、生活、生態的在地產業,卻也弄得像工業或高科技產品,農產品的強權到你們家來說你們不要打籃球了,因為你們打不過我們!身高不夠、技不如人,連娛樂健身的餘地都沒有,自給自足只是浪漫的脫節,真是無奈的所在!
被定義為敲門磚的ANZTEC
我國與紐西蘭的經濟合作協定(ANZTEC)已於7月10日簽署,以國家的立場來說算是喜事一件,但是看到ANZTEC的簽署,心情頗為複雜。幾年來國際間拼命在經貿上結盟,台灣遲遲未有進展,都快變成亞細亞的孤兒了,加上經濟一直難有起色,政府壓力之沉重可想而知,好不容易簽了第一個貿易協定,為什麼偏偏是紐西蘭呢?紐西蘭的強項你我皆知,哪些產業會是馬前卒猜也不用猜,會否有衝擊大家心知肚明。我國與紐西蘭貿易額雖不大,但被定義為敲門磚,政府說這是臺灣與主要貿易夥伴簽署的第
一份完整自由貿易協定,對我國經濟發展真有重大意義。根據中華經濟研究院的分析,國家整體產值可增加141∼356億元,而農業產值則將減少25∼35億元(喔,有不少農產業是「籃球」.
. . . )。政府又說:紐西蘭是跨太平洋戰略經濟夥伴關係協議 (TPP) 及區域全面經濟夥伴關係(
RCEP,主要目的為推動東協十國與中國等其它六國間的自由貿易)的重要成員,這次臺紐兩國簽署經濟合作協定,對於我國加入相關區域經濟整合,或是與其他國家簽署自由貿易協定,將有極大幫助,而我農業部門也可以利用這次機會,積極調整產業結構與提升競爭力,因應未來加入TPP的嚴峻挑戰(喔,至少還有機會練球,原來ANZTEC是預賽......)。國家是一體的,產業團體也充分爭取了,但形勢比人強,我們說
NO也沒有用,還好乳業掙得了一個12年調整的喘息空間,只能安慰人生之不如意十之八九,辛苦與挑戰從來不曾少過,也還好經過多年的努力,酪農業不會真的像是台灣打不起的籃球。
●前言
目前生乳收購計價根據的是乳脂率以及非乳脂固形物,國內酪農對於提高乳脂率的方法相當熟悉,甚至可以提高乳脂率超過生乳收購計價標準的乳脂率最高限4.0%,但是乳脂率超過4.
0%之後,乳品工廠並不會支付更高收購價格。反倒是能夠達到非乳脂固形物計價標準最高限的路農戶並不多。如果想要有更高的乳款收入,勢必要從調整非乳脂固形物下手才有最大空間。調整非乳脂固形物比較容易下手的切入點,主要包含飼養、疾病、牛群管理等三個方向。
●飼養
通常只要泌乳牛飼糧添加脂肪,就會造成乳蛋白質率下降,一般都可以使用下列公式預估乳蛋白質率下降程度:
添加脂肪組乳蛋白質率(%) /對照組乳蛋白質率(%)=101.1-0.6381X+0.0141X 2 ;X=飼糧脂肪(%)
套用上述公式,可以簡單推算出泌乳牛飼糧添加脂肪後乳蛋白質率下降程度如表1所示。而且要留意的另一件事情是泌乳牛飼糧添加脂肪經常會造成採食量下降,這對分娩前後過渡時期的乳牛特別不利。因此一般建議的脂肪添加時機為泌乳期40∼120天。而且要同時提高飼糧配方過瘤胃蛋白質或胺基酸比例,因為提高飼糧配方過瘤胃蛋白質或胺基酸比例,可以幫忙拉高乳蛋白質率(表2,Canale
et al.,1990)。
表1.泌乳牛飼糧添加脂肪後所造成相對於未添加脂肪時候的乳蛋白率降低程度
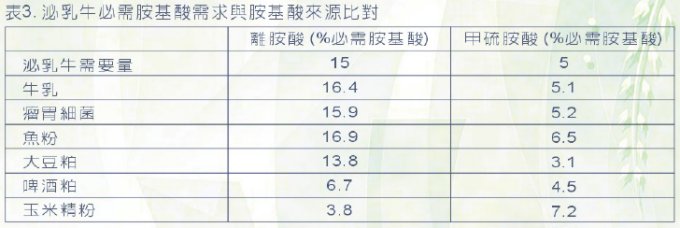
泌乳牛可消化必需胺基酸的需要量如表3所示 (Doepel et
al.,2004),通常飼糧中的總胺基酸的消化吸收約在70%,所以想推算回飼糧總總蛋白質中各個胺基酸的量,就將表3的數據除以0.7即可。事實上必需胺基酸組成最符合泌乳牛需求的是瘤胃細菌
(表4 ; Santos et al., 1998),通常的做法是先確認錯糧自己方提供定量的瘤胃可分解蛋白質與可利用非纖維碳水化合物
(例如熟化玉米),讓瘤胃細菌生長達到最佳狀態,然後才去檢討需要再額外添加多少過瘤胃蛋白質或胺基酸。
●疾病
酮症的泌乳牛會有較高的乳脂率,根據Stevens (2012)的研究是正常牛乳脂率的1.3倍。Duffield et
al.
(1997)檢討將乳蛋白質率/乳脂肪率小於或等於0.75、乳脂肪率大於或等於4.1%、乳蛋白質率小於或等於3.1%,共同當為可能發生酮症的預篩指標的敏感度與專一位仍不夠高。所以目前能確定的是,酮症泌乳牛的乳脂率會偏高,但不可以因為乳脂率偏高就斷言是銅症。要確診是不是酮症,仍然要從乳汁或尿液進行酮體檢測。分娩前到分娩後的過渡時期,每頭母牛投給400
mL的丙二醇,可以有預防酮症並且提高乳量的效果。當然,可以推測 少了酮症發生機率,相對的非乳脂固形物要維持較高的機會就較高。
熱緊迫會降低乳蛋白質率 (3.01 VS. 3.31%,Bernabucci et al.,
2002 ; 2.68 VS. 2.97%,Kamiya et al., 2006),也會降低乳量,Correa-Calderon et al. (2006)
試驗發現使用降溫系統可以提高泌乳牛的乳量 (32.7 VS. 28.5 公斤/日),但是沒有同時提高乳蛋白質率 (2.85 VS.
3.00%)。可能是營養份被轉換去合成所有乳成份,而不是專一使用於合成乳蛋白質。因此,使用降溫系統的同時,也要同時留意飼糧針對熱緊迫應有的調整,才不會功效不如期望。熱緊迫應有的銷糧調整包含(1)只餵高品質牧草,盡量降低飼糧纖維含量(中洗纖維
25-30%,酸洗纖維14-17%),以降低發酵熱負擔;(2)短纖維副產物敢代最多1/3的芻料;(3)調高過瘤胃蛋白質達40%粗蛋白質;(4)調高非纖維碳水化合物
(35%乾物質)或脂肪(7%乾物質)含量;(5)調高礦鹽,Na 0.45-0.6%、K 1.5-1.6%、CI0.25%、Mg0.35-0.4% ;
(6)添加與免疫、繁殖相關維生素:維生素A 100000-200000 IU/日/頭、維生素E10001U/日/頭。
乳房炎會造成乳脂肪率下降程度大於乳蛋白質率下降程度
(Harmon,1994),生乳體細胞數跟乳蛋白質率/乳脂肪率有顯著性的正相關 (Jamrozik and
Schaeffer,2012),因此只有在正常體細胞數範圍內看到乳蛋白質率/乳脂肪率比例升高才是值得高興的事。預防乳房炎,可以從乾乳期乳房炎要高處理以及乳頭封膠的使用之外,還要注重分娩前到分娩後的飼糧維生素E(乾乳牛的1000
IU/日;泌乳牛5001U/日)、硒 (0.3 ppm)、銅 (20 ppm)、鋅 (40-60 ppm) 等養分的足量供應。
● 牛群管理
乳蛋白質率/乳脂肪率比例會隨著泌乳天數增加而提高 (Jamrozik
and
Schaeffer,2012),所以只要處於泌乳初期的泌乳牛在牛群中的比例不過半,整個泌乳牛群的總乳乳蛋白質率/乳脂肪率比例就能維持穩定狀態。另一個影響乳蛋白質率/乳脂肪率比例是泌乳期,從第一泌乳期到第五泌乳期,每年乳脂肪率下降0.2%單位,乳蛋白率下降0.02-0.05%單位,隨著胎次增加,乳蛋白質率/乳脂肪率比例會提高。因此,只要提高泌乳牛群的使用壽命,自然可以提高泌乳牛群的總乳乳蛋白質率/乳脂肪率比例,這代表經營成本下降,經營效率提高,
而且可以獲得高非乳脂固形物的高收購乳價。不同乳牛品種有不同的乳組成,如果同時有飼養荷蘭牛與娟姍牛,可以籍由調整荷蘭牛與娟姍牛的比例,就可以調整出不同的總乳的乳脂肪率以及非乳脂固形物(表4)。如果只有養一種荷蘭牛也沒關係,因為乳組成具備有中度的遺傳率,只要選留高非乳脂固形物的母牛,後面繁衍出來的新牛群就會有較高的非乳脂固形物表現。
● 結語
只要我們清楚了解飼糧飼糧脂肪、瘤胃可分解蛋白質、瘤胃可利用非纖維碳水化合物等成份的調節,要同時考慮過瘤胃蛋白質或胺基酸的調整。避免酮症與熱緊迫的對泌乳牛群侵擾,同時從各方面努力延長泌乳牛使用壽命,再加上善用乳組成遺傳率效應,整體的努力一定可以有效提高非乳脂固形物,增加酪農的乳款收入。
●參考文獻
Bernabucci, U., N. Lacetera, B. Ronchi, and A.
Nardone. 2002. Effects of the hot season on milk protein fractions in Holstein
cows. Anim.
Res. 51:2533.
Canale, C. J., L. D. Muller, H. A. McCahon, T. J. Whitsel, G. A. Varga and M. J.
Lormore. 1990. Dietary fat and ruminally protected
amino acids for high producing
dairy cows. J. Dairy Sci. 73:135-141.
Correa-Calderon, A., D. V. Armstrong,D. E. Ray,S. K. DeNise,R. M. Enns and C. .
Howison. 2005. Producr\tive and reproductive
response of Holstein and Brown
Swiss heat stressed dairy cows to two different cooling systems. J. Anim. Vet.
Adv. 4:572-578.
Doepel, L., D. Pacheco, J. J. Kennelly, M. D. Hanigan, 1. F. Lopez and H.
Lapierre. 2004. Milk protein synthesis as a function of amino
acid supply. J. Dairy Sci.
87:1279-1297.
Duffield, T. F., D. F. Kelton, K. E. Leslie, K. D. Lissemore, and J. H. Lumsden.
1997. Use of test day milk fat and milk protein to detect
subclinical ketosis in dairy cattle
in Ontario. Can. Vet. J. 38: 713-718.
Harmon R. J. 1994. Physiology of mastitis and factors affecting somatic cell
counts. J Dairy Sci. 77:2103-2112.
Jamrozik, J. and L. R. Schaeffer. 2012. Test-day somatic cell score,
fat-to-protein ratio and milk yield as indicator traits for sub-clinical
mastitis in dairy cattle. J. Anim. Breed.
Genet. 129: 1119.
Kamiya, M., Y. Kamiya, M. Tanaka and S. Shioya1. 2006. Milk protein production
and plasma 3-methylhistidine concentration in
lactating Holstein cows exposed to high
ambient temperatures. Asian-Aust. J. Anim. Sci. 19:1159 1163.
Santos, F. A. P., J. E. P. Santos, C. B. Theurer and J. T. Huber. 1998. Effects
of rumen-undegradable protein on dairy cow performance: a
12-year literature review. J. Dairy Sci. 81:
3182-3213.
Stevens, K. D. 2012. Monitoring the incidence of ketosis in fresh cows using
milk composition, urine ketones, and milk ketones. Honors
Research Thesis,Ohio State Univ.
[回上頁 | 顯示此文件為可列印格式]
此文件提供者: shuYing - [評分 : 0.00 (0 票選) | 評分!]
|
|
| 所徽 |
|

|
| 隨機小語 |
出類拔萃的人都有一個共通點:使命感。
[金克拉] |
|